


Research Progress on Bacteriocins Produced by Lactic Acid Bacteria and Their Inhibitory Effect on the Biofilm of Staphylococcus aureus
-
摘要: 金黄色葡萄球菌(Staphylococcus aureus,S. aureus)是一种能在食品表面形成生物膜的革兰氏阳性条件致病菌,其生物膜的形成对食品加工和食品安全造成了严重威胁。乳酸菌细菌素是由乳酸菌在代谢过程中产生的一类具有抗菌活性的多肽,可抑制多种食源性致病菌及其生物膜的形成。本文针对乳酸菌细菌素的分类、作用方式及其在群体感应调控下的合成机制进行了综述,基于培养条件和培养基成分简述了提高乳酸菌细菌素合成量的常规方法,总结了不同来源乳酸菌细菌素的纯化方式,系统综述了乳酸菌细菌素对S. aureus生物膜的抑制作用,从Agr系统和LuxS/AI-2群体感应系统的角度简要阐述了乳酸菌细菌素对S. aureus生物膜的调控机制,并展望了乳酸菌细菌素对S. aureus生物膜抑制机制的未来研究方向,为乳酸菌细菌素作为生物防腐剂的开发和广泛使用提供科学依据。Abstract: Staphylococcus aureus (S. aureus) is a Gram-positive opportunistic pathogen that can form biofilm on the surface of food. Its biofilm formation poses a serious threat to food processing and food safety. Bacteriocins are peptides with antimicrobial activity produced by lactic acid bacteria (LAB) during metabolism. They can inhibit a variety of food-borne pathogens and the formation of biofilm. In this paper, the classification, mode of action, and synthesis mechanism under the regulation of quorum sensing of LAB bacteriocins are summarized. Furthermore, the conventional methods to improve the synthesis of LAB bacteriocins are briefly described based on the culture conditions and media components. The inhibition of LAB bacteriocins on the biofilm of S. aureus is systemically reviewed. Moreover, with particular reference to the Agr and LuxS/AI-2 quorum sensing systems, the regulatory mechanisms of S. aureus biofilm formation by LAB bacteriocins are briefly outlined. In addition, the future research direction on the mechanism of inhibition of S. aureus biofilm by LAB bacteriocins is prospected. This is expected to provide a scientific basis for the development and widespread application of LAB bacteriocins as biopreservative agents.
-
金黄色葡萄球菌(Staphylococcus aureus,S. aureus)是典型的食源性致病菌,可产生肠毒素、溶血素和杀白细胞素等多种毒力因子[1],且极易附着在乳制品和肉类等各种食品加工设备表面并形成生物膜[2],导致食品中潜在的细菌交叉污染。生物膜是嵌入在由多糖、蛋白质和细胞外DNA(Extracellular DNA,eDNA)组成的胞外基质中的一种或多种微生物的聚集体,有助于增强细菌毒性和对抗生素的抗性[3]。
细菌素是一种由核糖体合成的有抗菌活性的蛋白或多肽类物质[4],可抑制多种食源性致病菌。乳酸菌产生的细菌素因其安全、高效的特点在食品领域有着广阔的应用前景,受到了越来越多研究者的关注[5]。乳酸菌细菌素可通过群体感应(Quorum sensing,QS)的方式调控S. aureus生物膜的形成,并通过靶向细胞膜、孔形成、质子动力损失、肽聚糖合成抑制和细胞质渗漏等方式抑制S. aureus生物膜的形成[6],有望成为传统抗生素和化学防腐剂的天然替代品。
本文简述了乳酸菌细菌素的分类、作用机制及群体感应调控下Autoinducing peptide(AIP)和Autoinducer-2(AI-2)介导的细菌素合成机制,并对细菌素的分离纯化过程进行了详细总结;重点综述了乳酸菌细菌素对S. aureus生物膜的抑制机制,为开发更多可抑制食源性致病菌的细菌素提供参考。
1. 乳酸菌细菌素的分类及抑菌机制
1.1 乳酸菌细菌素定义
乳酸菌细菌素是在乳酸菌代谢过程中通过核糖体合成的一类抗菌蛋白或多肽,可以有效抑制病原微生物生长。一种细菌产生的细菌素不但可以对该菌的同种或近缘生物起到抑制作用,对其他多种腐败微生物或食源性致病菌也可能起到抑制作用[4]。大部分细菌素是由食品级的乳酸菌产生的,乳酸菌是公认的安全菌株,被广泛应用于食品、生物和医疗等方面[7]。大部分乳酸菌细菌素对热稳定,在大多数食品体系中的宽pH范围内具有稳定的生物活性,无毒,无残留,无耐药性,不影响食品感官质量,并对一系列食品致病菌和腐败菌具有抗菌活性[8]。此外,乳酸菌细菌素对胰酶复合物、胰蛋白酶和胰凝乳蛋白酶等消化蛋白酶敏感,不会对肠道微生物群产生负面影响[9]。因此,乳酸菌细菌素在食品工业有着极大的应用前景,有望替代传统的化学防腐剂,一直是食品领域关注的焦点。
1.2 乳酸菌细菌素分类
乳酸菌细菌素可根据其物理性质、化学结构、稳定性、作用方式、分子大小以及抗菌活性等进行分类[10−11]。为更好地研究乳酸菌细菌素,多数研究者按照其分子结构特征将其分为3类:Class I、Class Ⅱ和Class Ⅲ。具体分类见表1。
表 1 细菌素分类Table 1. Classification of bacteriocinsClass I是由某些特殊氨基酸组成的细菌素,如羊毛硫氨酸、甲基羊毛硫氨酸、脱氢丁酸和脱氢丙氨酸,所以又被称为羊毛硫抗生素,Nisin是Ⅰ类细菌素的典型代表[12]。Class Ⅱ是一类未经修饰或极少修饰的细菌素,它们的成熟不需要除前导肽酶和转运蛋白以外的酶,Class Ⅱ的稳定性和抑菌活性更强,抑菌谱更广。Enriqueta等[13]将Class Ⅱ细分为四个亚类:Class IIa、Class IIb、Class IIc和Class IId。Class IIa也被称为类片球菌素,对多种食源性致病菌如芽孢杆菌、梭状芽孢杆菌、葡萄球菌等均有抑制作用;Class IIb是双肽细菌素,由两种物质的量相等的肽组成[14],两种肽相互作用以发挥其最佳的抗菌活性;Class IIc是环状细菌素,其特征是有一个肽键连接核心肽的N端和C端;Class IId是无前导序列的细菌素[15]。Class Ⅲ是未修饰细菌素,具有溶菌或非溶菌作用机制[16]。
1.3 乳酸菌细菌素抑菌机制
乳酸菌细菌素的主要作用方式是破坏指示菌的细胞膜,使细胞膜形成孔隙并被裂解,从而使无机磷酸盐等细胞内容物流出,导致细胞死亡[17]。如plantaricin FB-2[18]通过破坏S. aureus细胞膜结构,增加细胞通透性,促进细胞质内容物(如蛋白质、DNA、K+、Ca2+等)渗漏,增加细胞内活性氧水平而发挥抗菌作用;Plantaricin Z057[19]可破坏副溶血弧菌(Vibrio parahaemolyticus)细胞膜完整性,导致细胞变形和细胞质渗漏,从而导致细菌细胞死亡。除此之外,细菌素还可以通过破坏细胞壁、抑制细胞DNA合成和氨基酸合成等方式作用于致病菌[20],如细菌素Y19-2通过破坏细胞壁和影响DNA及RNA的合成来抑制S. aureus和大肠杆菌(Escherichia coli, E. coli)的生长[21],细菌素BM1157可以抑制S. aureus细胞DNA合成[22]。
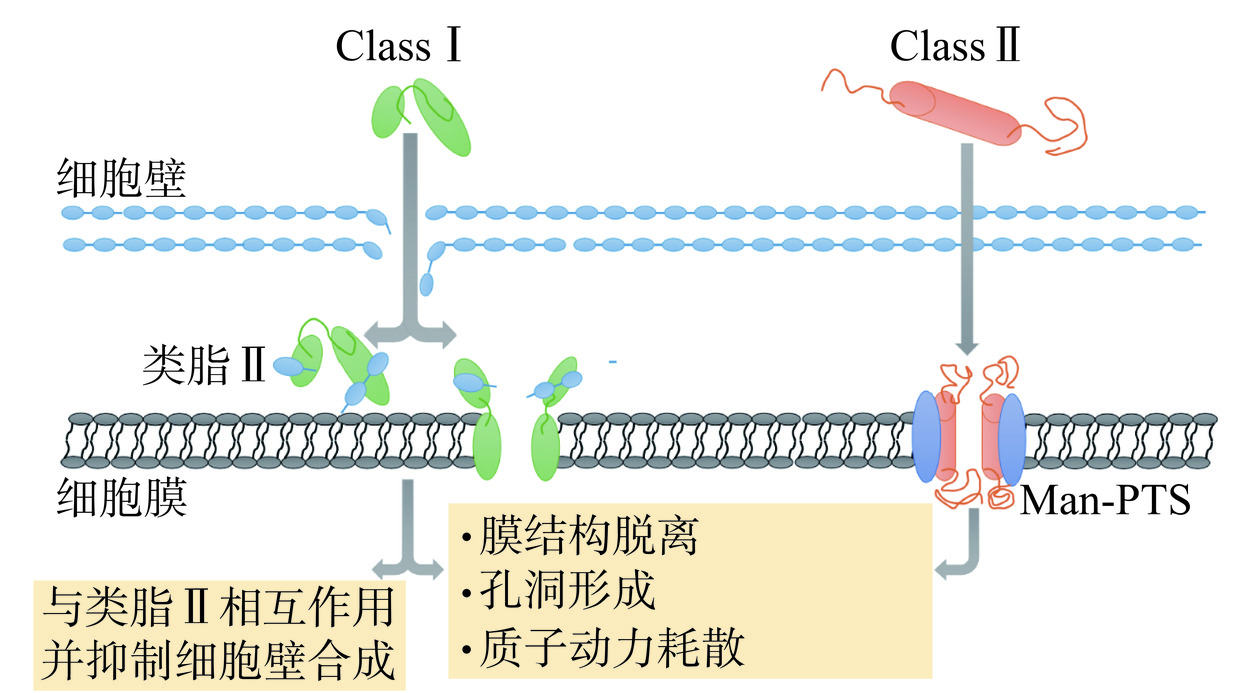
不同结构的细菌素,其作用方式也有所不同。Class Ⅰ细菌素主要是羊毛硫细菌素(如Nisin),它能以高亲和力与细胞壁合成的前体分子类脂II结合来抑制细胞壁合成[23](图1)。焦磷酸基团在细胞壁合成中起关键作用,类脂II分子的部分焦磷酸基团与Nisin N-端的氨基化合物之间形成5个分子间氢键,可以抑制细胞壁的形成[24]。此外,Nisin还可以通过在细菌细胞膜上形成孔来发挥作用。Nisin的氨基末端与类脂II分子结合,抑制肽聚糖和羧基末端的合成,导致细菌细胞膜上形成孔隙,使细胞内容物流出,抑制细菌生长[25]。
Class Ⅱ细菌素的作用方式与Class Ⅰ相似,通过破坏细菌细胞膜导致细胞死亡[26]。Ⅱa类细菌素(如pediocin)与甘露糖磷酸转移酶系统(Mannose Phosphotransferase System, Man-PTS)结合,使细胞膜形成孔隙,破坏电势(Δψ)和跨膜质子梯度(ΔpH)中的一种或两种来耗散质子动力势(PMF),导致靶细胞死亡[6]。例如Pediocin PA-1与Man-PTS结合可使单增李斯特菌(Listeria monocytogenes)细胞膜上形成孔洞,耗散PMF,从而达到抑菌效果[27];Pediocin L50可使E. coli和幽门螺旋杆菌(Helicobacter pylori)细胞膜表面形成孔隙,最终导致细菌死亡[28]。IIb与IIc类细菌素的抑菌作用均是基于靶细胞膜的通透性,使PMF耗散和细胞内信息分子泄露[15]。Lacticin Q是典型的IId类细菌素,其抑菌机制是直接与细菌细胞膜相互作用,从而引起渗透,导致离子泄漏、膜电位耗散和细胞死亡[29]。
2. 群体感应调控下的乳酸菌细菌素合成机制
乳酸菌细菌素的合成可能涉及不同的基因和蛋白质,所以其合成模式也不尽相同,调控其合成的机制主要有QS、环境诱导、SOS应激等,其中以QS调控细菌素生物合成的研究较多[30]。QS是革兰氏阳性菌和革兰氏阴性菌在应对环境变化时调节细菌行为和基因表达的一种调控机制[31]。在细胞密度达到阈值时,细菌会产生并释放被称为信息素或自诱导剂的信号分子,从而启动QS调控细菌素合成,杀死目标细菌[32]。调控乳酸菌细菌素合成的群体感应系统主要有两类:Autoinducing peptide(AIP)介导的群体感应系统(种内调控系统)和Autoinducer-2(AI-2)介导的LuxS/AI-2群体感应系统(种间调控系统)[33]。
2.1 AIP诱导下的乳酸菌细菌素合成
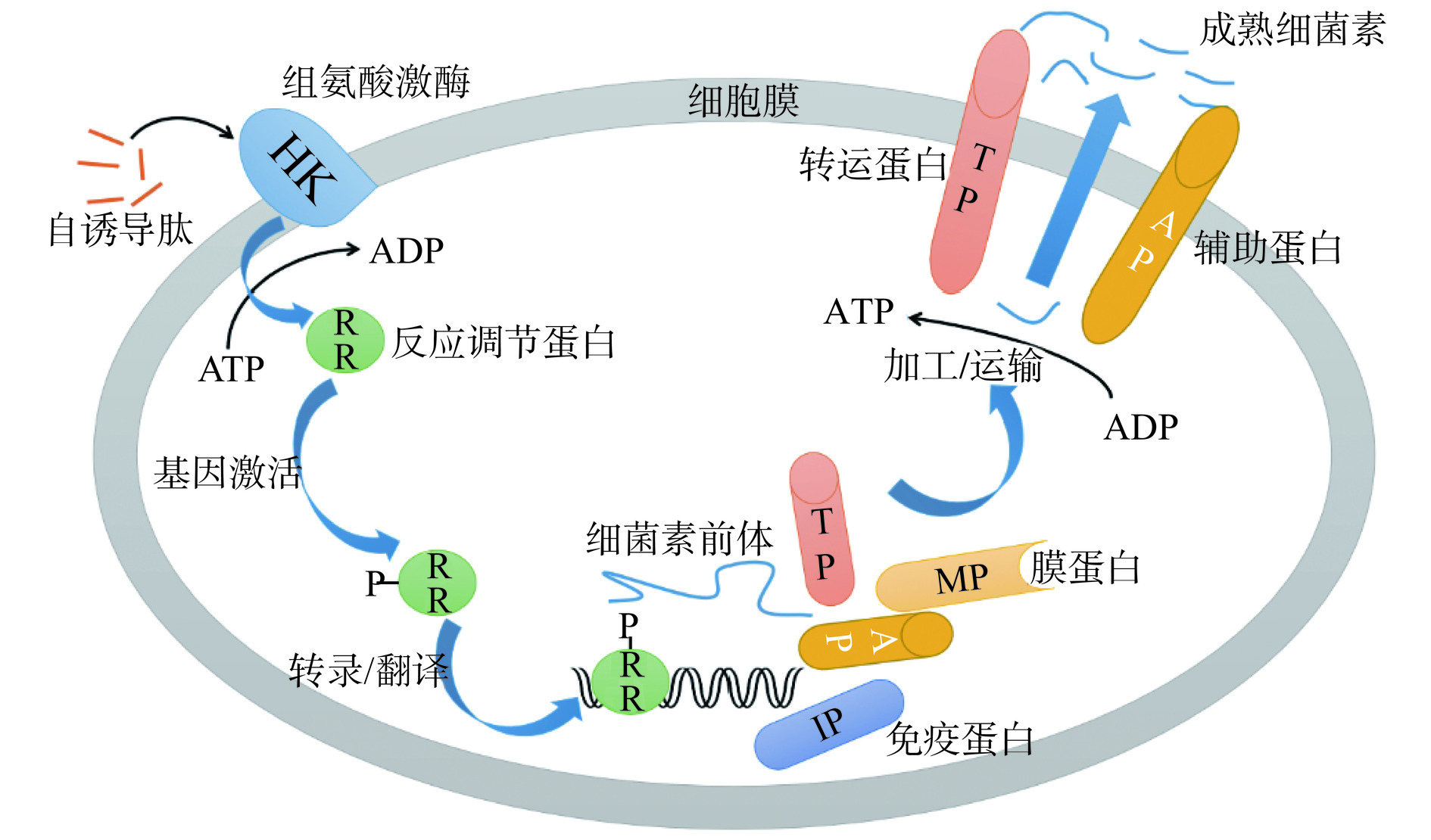
自诱导肽AIP是种内QS系统的信号分子,在核糖体中合成后经过修饰加工达到成熟形式[34],不同种类细菌合成的AIP也不同,具有高特异性[35]。AIP随乳酸菌的增殖而不断合成并被转运至胞外,当其浓度积累到一定程度时,可通过细胞膜上的敏感结构域激发组氨酸蛋白激酶(Histidine protein kinase,HPK)的自磷酸化,并进一步使细胞质中的反应调节蛋白(Response-regulating protein,RR)磷酸化。随后,磷酸化的RR与目标功能基因的启动子结合,诱导其表达,进而开始合成并分泌细菌素。最后,这些细菌素可以通过ABC转运体在细胞外释放,成为具有抑菌活性的成熟细菌素[36],如图2。植物乳杆菌(Lactobacillus plantarum, L. plantarum)NC8[35]和清酒乳杆菌(Lactobacillus sakei)Lb706[37]产细菌素都是以AIP作为信号分子,还有一些细菌素如Nisin和subtilin以其自身作为AIP来激发信号转导,以诱导其自身的合成[38]。
![]() 图 2 群体感应调控下的细菌素合成与转运机制Figure 2. Mechanism of bacteriocins synthesis and transport regulated by quorum sensing
图 2 群体感应调控下的细菌素合成与转运机制Figure 2. Mechanism of bacteriocins synthesis and transport regulated by quorum sensing2.2 AI-2诱导下的乳酸菌细菌素合成
AI-2是一种呋喃酮化合物,在种间细胞交流中起着信号分子的作用,由细菌中的LuxS蛋白催化合成[39]。LuxS是一种代谢酶,参与细菌硫代谢的甲基循环,催化S-核糖高半胱氨酸(S-ribosylhomocysteine,SRH)转化为高半胱氨酸和4,5-二羟基-2,3-戊二铜(4,5-Dihydroxy-2,3-pentanedione,DPD),DPD进一步合成AI-2[40]。在高细胞密度下,AI-2与受体蛋白(LuxP)结合后,再与传感器激酶(LuxQ)相互作用,从而引发磷酸化反应,诱导生物发光基因和luxS基因的转录[41]。LuxS基因是编码LuxS蛋白的基因,在许多乳酸菌菌株中,luxS基因转录水平与AI-2活性呈正相关[38]。目前虽没有研究明确表明细菌素的产生与AI-2或luxS基因有直接关系,但一些研究者通过乳酸菌共培养发现AI-2对细菌素的产生有正向调控作用,如Jia等[42]发现缺失luxS基因的L. plantarum KLDS1.0391与正常的瑞士乳杆菌(Lactobacillus suisse)KLDS1.9207共培养时,在4~24 h的生长期内,其AI-2活性均显著低于正常菌株(P<0.05),且L. plantarum KLDS1.0391的细菌素产量与AI-2活性呈正相关,该结论与Liu等[43]研究的L. plantarum RX-8与枯草芽孢杆菌(Bacillus subtilis)BS-15共培养所得结论一致,表明luxS基因在共培养过程中对细菌素的产生有较大影响;L. plantarum NMD-17与罗伊氏乳杆菌(Lactobacillus reuteri)NMD-86共培养时,其细菌素合成基因plnB、plnD、luxS、plnE和plnF的转录水平显著上调(P<0.05),AI-2活性显著提高(P<0.05),表明L. plantarum NMD-17在共培养中的细菌素合成可能与AI-2介导的群体感应系统密切相关[44]。Li等[45]将L. plantarum AB-1和干酪乳酪杆菌(Lacticaseibacillus casei, L. casei)共培养,与L. plantarum AB-1单独培养相比,其抗菌活性显著提高,群体感应基因luxS和细菌素调节操纵子(PlnB和PlnC)的转录也显著增加,将其共同接种到对虾中,对虾中的腐败菌受到显著抑制,AI-2活性显著升高,表明LuxS/AI-2系统可以调节乳酸菌的协同作用和细菌素的产生。
3. 乳酸菌细菌素产量优化
迄今为止,商品化的细菌素相对较少,其原因之一是细菌素产量低,提高细菌素的产量是目前亟待解决的问题。乳酸菌细菌素的产生与菌株所处的外界环境密切相关,培养基初始pH、温度、接种浓度、发酵时间等培养条件都对细菌素的合成量有一定影响[46−47]。近年来一些研究者通过对乳酸菌产细菌素条件的优化使细菌素产量得以提高。如黄倩等[48]通过响应面试验对乳酸乳球菌(Lactobacillus lactis, L. lactis)Q13的发酵条件进行优化,得到最优发酵条件为接种量1.4%(v/v)、发酵初始pH8.0、发酵温度27.5 ℃,此条件下,细菌素效价比优化前提高3倍;唐美[49]通过单因素实验、Plackett-Burman试验和响应面试验优化卷曲乳杆菌(Lactobacillus crispatus)产抑菌物质的发酵条件,得到最佳发酵条件为初始pH8.1、培养时间32 h、接种量为4%(v/v)、培养温度37 ℃,优化后抑菌物质产量提高42.2%;Jawan等[50]通过研究培养条件对L. lactis Gh1产生类细菌素抑制物质的影响,发现接种浓度为1%(v/v)时细菌素产量最高。
除培养条件外,培养基成分对细菌素合成量也有着重要影响。碳源能增加酶和代谢化合物的生成,促进生物量的产生,从而提高细菌素的合成量,如Ye等[51]已通过基因表达分析和结构方程建模证明了适宜的碳源浓度和碳源类型可以增加副干酪乳酪杆菌(Lacticaseibacillus paracasei, L. paracasei)HD1.7细菌素的合成量。氮源具有多种氨基酸和多肽来促进细胞生长并诱导细菌素的产生;此外,吐温80、碳水化合物和盐浓度等也对细菌素的产量有一定影响[50,52]。Parlindungan等[53]的研究表明氮源及吐温80对L. plantarum B21细菌素的合成至关重要。不同细菌产细菌素所需要的营养成分也不同,因此近年来一些研究者通过对培养基成分进行优化来提高细菌素的合成量和活性,例如Daba等[5]使用Plackett-Burman设计和Central Composite设计对MRS培养基成分进行了优化,与未优化前相比,使用优化培养基的细菌素产量从800 AU/mL增加到12800 AU/mL。Li等[54]使用与响应面试验相关的析因设计方法研究了培养基中六种不同营养成分对细菌素产生的影响,并优化得到最佳培养基成分组成,与常规培养基相比,优化后的培养基可使细菌素产量增加1倍。因此,采用一些优化方法如析因设计和Plackett-Burman设计等对培养条件和培养基成分进行优化可有效提高细菌素产量,促进细菌素工业化生产。
4. 乳酸菌细菌素分离纯化
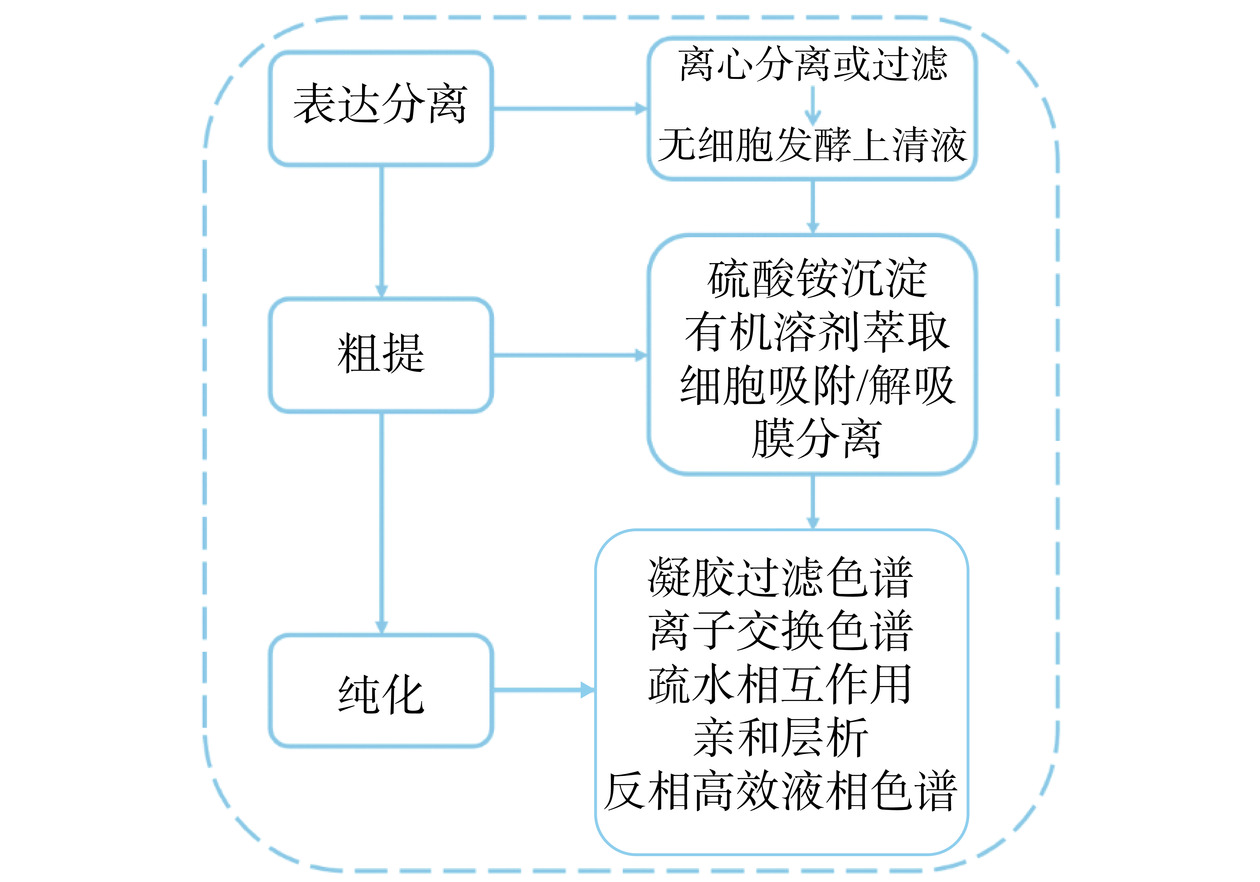
细菌素粗提、纯化的一般步骤如图3所示。
4.1 乳酸菌细菌素粗提
细菌素一般产生于乳酸菌发酵过程中,主要溶解在发酵液中,并且含量较低,所以需要人工进行粗提,其目的是得到澄清、稳定的粗蛋白,为理化性质的研究及后续蛋白纯化奠定基础。实验室中常用的粗提方法有硫酸铵沉淀法、有机溶剂萃取法、细胞吸附/解吸法和膜分离技术等,如表2所示。硫酸铵沉淀法通常需要分级沉淀才能得到纯净的提取物,在沉淀过程中会引入盐离子并需在后续的纯化过程中去除。有机溶剂萃取则相对简单,无需脱盐。细胞吸附/解吸法可以根据目标蛋白的吸附难易程度选择不同的吸附方式,较为灵活。膜分离技术则比较适合大规模工业生产。
表 2 细菌素粗提方法Table 2. Crude extraction methods of bacteriocins方法 原理 特点 文献 硫酸铵沉淀法 高浓度盐破坏蛋白质分子表面的水化层,与蛋白质竞争水分子,使蛋白质溶解度降低并形成沉淀,从而从溶液中
分离出来优点:溶解度高、成本低、操作简单、蛋白质在
高浓度盐条件下不易变性
缺点:粗提产物含大量杂蛋白,后期纯化难度大[55] 有机溶剂萃取法 基于细菌素的极性,将有机溶剂与细菌素混合,利用相似相溶原理将细菌素从水相溶解进入有机相,从而实现
细菌素的浓缩优点:有机溶剂易于除去
缺点:成本高、有机溶剂后处理时对环境的
污染问题[56−57] 细胞吸附/解吸法 利用细菌素的带电特性,调节溶液的pH可使菌体细胞吸附细菌素,再调回溶液原来的pH便可使细菌素解吸 优点:成本低和利于后期进一步纯化
缺点:有一定局限性,不具有普遍性[58] 膜分离技术 依据细菌素的分子量大小,选用合适的膜进行截留,达到分离的目的 优点:高选择性、不需要添加剂、易于扩大规模
缺点:样品损失量多,不易清洁[59] 4.2 乳酸菌细菌素的纯化
乳酸菌细菌素进行初步粗提后,一般需要进一步的纯化以得到纯度更高的细菌素。通常用色谱法进行纯化,如凝胶过滤色谱、离子交换色谱、疏水相互作用、亲和层析和反相高效液相色谱等。
凝胶过滤色谱是根据不同分子量的蛋白质分子在凝胶柱内的流速不同来进行分离,一般用于除去粗提样品中的小分子的杂蛋白和盐析过程中残留的硫酸铵或氯化钠[60];由于细菌素分子在不同的pH下带有不同的电荷,所以离子交换色谱法也常用来纯化细菌素,离子交换色谱法是一种灵活的方法,可以通过选择最合适的离子交换器、控制pH和缓冲液离子强度来进行优化,由于乳酸菌细菌素大多都带正电荷,所以通常用阳离子交换层析来分离乳酸菌细菌素[61];当细菌素高度疏水时,可用疏水相互作用进行分离,该方法是基于不同分子之间疏水性差异不同,从而导致与固定相之间作用强弱的不同来分离样品[62]。此外,以细菌素的特异性抗体为配体的亲和层析也常用于纯化细菌素,该方法选择性强、分离效果好且操作时间短;反相高效液相色谱法(Reversed-phase high-performance liquid chromatography,RP-HPLC)常用于整个纯化过程的最后一步,其原理是基于不同的溶质分子与固定相疏水基间的疏水相互作用不同来进行分离。通过该方法分离纯化后,细菌素纯度可达到99%以上,可用于氨基酸序列和质谱分析[63]。
乳酸菌细菌素因其性质不完全相同,故所产细菌素需要不同的纯化方法。如L. plantarum SLG10所产的细菌素SLG10可用生物色谱法与RP-HPLC相结合的方法来进行分离纯化[64],而鼠李糖乳杆菌(Lactobacillus rhamnosus,L. rhamnosus)LS-8细菌素的纯化过程则相对复杂,需要通过硫酸铵沉淀、细胞吸附-解吸、阳离子交换色谱和RP-HPLC等方法进行分离纯化[65]。表3总结了近几年来不同来源菌株所产细菌素的主要纯化方法。
表 3 细菌素纯化常规方法Table 3. Bacteriocins purification conventional method纯化方法 微生物菌株及来源 细菌素 分子量 文献 疏水相互作用色谱、固相萃取和RP-HPLC 发酵乳杆菌(Lactobacillus
Fermentum, L. Fermentum)GA715,羊乳Fermencin SA715 1792.537 Da [52] 凝胶过滤色谱和HPLC L. casei LiN333,江水菜 细菌素LiN333 4890 Da [66] 硫酸铵沉淀、pH介导的细胞吸附-解吸、阳离子交换色谱和RP-HPLC L. rhamnosus LS-8,四川泡菜 细菌素pH 25、S68、S81和S137 / [65] 硫酸铵沉淀、阳离子交换色谱和RP-HPLC L. plantarumSHY 21-2,牦牛酸奶 Plantaricin LP 21-2 1362.84 Da [67] 乙酸乙酯萃取、阴离子交换色谱和HPLC L. rhamnosus zrx01,Kefir 细菌素zrx01 / [68] 阳离子交换色谱和疏水相互作用色谱 L. plantarum GCNRC_GA15,埃及山羊奶酪 细菌素GA15 4369 Da [5] Superdex 30 Increase 10/300 GL排阻色谱 L. paracasei LS-6,发酵酸奶 细菌素LSX01 967.49 Da [69] 大孔树脂XAD-16吸附色谱、阳离子交换色谱、Sephadex G-25凝胶过滤色谱和RP-HPLC L. paracasei ZFM54,新生婴儿粪便 细菌素ZFM54 1143.39 Da [70] 乙酸乙酯萃取和阴离子交换色谱 L. rhamnosus 1.0320,酸马奶 细菌素1.0320 1~3.3 kDa [57] Superdex™ 30 Increase 10/300 GL排阻色谱 唾液乳杆菌(Lactobacillus salivarius, L. salivarius)CGMCC2070,云南黑骨鸡肠道粘膜 细菌素XJS01 666.31 Da [71] Ni-NTA亲和层析和RP-HPLC 结壳乳杆菌(Lactobacillus crustorum, L. crustorum)MN047,发酵马奶 细菌素BM173 / [72] 生物色谱法和RP-HPLC L. plantarum SLG10,康普茶 细菌素SLG10 1422 Da [64] 硫酸铵沉淀 L. plantarum PKLP5,生牛奶 细菌素Bac23 5.1 kDa [73] 乙酸乙酯萃取、Sephadex LH-20凝胶过滤色谱、阳离子交换色谱和RP-HPLC纯化 戊糖片球菌(Pediococcus pentosaceus)zy–B,
华贵栉孔扇贝肠道细菌素PE-ZYB1 2019.22 Da [56] 硫酸铵沉淀、阳离子交换色谱和RP-HPLC L. Fermentum BZ532,发酵谷物饮料 细菌素LF-BZ532 1105.563 Da [74] 双水相系统、Sephadex G - 50凝胶过滤色谱和RP-HPLC L. plantarum M1-UVs300,发酵香肠 细菌素M1-UVs300 3.4 kDa [75] 硫酸铵沉淀、固相萃取、阳离子交换色谱和RP - HPLC 屎肠球菌(Enterococcus faecium)HDX-2,腌渍黄瓜 肠道菌素HDX-2 5482 Da [76] 磁性脂质体吸附和RP-HPLC L. plantarum SLG1,牦牛干酪 Plantaricin SLG1 1083.25 Da [77] 硫酸铵沉淀和凝胶过滤色谱 L. lactis CH3,发酵乳制品 细菌素CH3 3.5 kDa [78] 5. 乳酸菌细菌素对金黄色葡萄球菌生物膜的抑制机制
5.1 金黄色葡萄球菌及其生物膜对食品工业的影响
生物膜是指附着在生命体或非生命体表面,并被胞外基质(多糖、脂质、蛋白质和eDNA)包裹的大菌落生物分子层[79],可为膜内细菌提供保护,增强对抗菌剂、清洁剂等的抗性。S. aureus是一种常见的食源性致病菌,极易在食品加工设备和食品基质表面形成生物膜,即便通过强的物理方法(机械冲洗、热蒸汽、超声波处理)和化学方法(清洁剂和消毒剂,如次氯酸钠、过氧化氢、过氧乙酸)处理也难以彻底将其清除[80]。S. aureus生物膜的形成会增强细菌毒性,进一步污染食品基质,导致食品反复交叉污染[81],不利于食品加工、流通与贮藏。在食品工业中,S. aureus生物膜污染主要分布在三个方面:a.食品原料,包括原料奶[82]、生肉[83]和海鲜[84]等食品中经常能分离出可形成生物膜的S. aureus,在适当条件下,S. aureus会黏附到食品表面,引起生物膜的初步形成。如Wang等[85]从原料奶中分离出S. aureus并发现该菌株能形成生物膜,生物膜的形成会加速原料奶的腐败变质;Wang等[86]从生鸡肉中分离出了可形成生物膜的S. aureus菌株,该菌株引起的污染缩短了生鸡肉货架期;Beshiru等[87]从即食虾中分离出S. aureus,结晶紫染色结果表明该菌株有生物膜形成能力,不利于即食虾的保鲜。b.食品加工环境和设备,包括运输管道、肉类屠宰场甚至操作人员的身体和衣服,都为S. aureus提供了绝佳的黏附场所,容易引起食品交叉污染[88]。c.流通过程,在食品储存、运输和销售过程中,储存环境不卫生、运输操作不当或销售人员操作不规范,都可能引起生物膜的形成[89]。S. aureus生物膜的形成会导致设备磨损、产品货架期缩短,给企业带来巨大经济损失[6]。因此,开发能够有效抑制生物膜形成而没有副作用的新型生物防腐剂至关重要。
5.2 乳酸菌细菌素对金黄色葡萄球菌生物膜的抑制机制
5.2.1 乳酸菌细菌素对金黄色葡萄球菌生物膜的抑制作用
S. aureus生物膜的形成是一个动态、循环的过程,其形成过程可分为以下四个阶段:粘附、聚集、成熟和分散。乳酸菌细菌素作为天然抗菌剂已被分离、表征和广泛研究,众所周知,Nisin有抗S. aureus生物膜活性[90],近年来也发现许多新型乳酸菌细菌素可抑制S. aureus生物膜的形成,如Krishnamoorthi等[78]通过结晶紫染色法测得纯化自L. lactis CH3的细菌素对S. aureus生物膜的抑制率可达到93.81%±0.66%,抑菌效果显著(P<0.05),通过扫描电镜观察到细菌素可使金黄色葡萄球菌细胞膜形成孔隙,并因此导致ATP、DNA、离子、酶和氨基酸等内容物流出;Qiao等[72]发现产自L. crustorum MN047的细菌素BM173可使S. aureus生物膜相对形成百分比从80.1%(1/32×MIC)降低到44.9%(1/2×MIC);Zhao等[91]研究表明plantaricin 827可下调S. aureus生物膜形成相关基因(如sdrC、sdrD、sdrE、arcA和sarA)的转录,抑制生物膜形成;Kim等[92]研究发现分离自乳酸片球菌(Pediococcus acidilactici)JM01的细菌素JM01可减少S. aureus浮游菌数量、降低黏附作用并显著降低生物膜内细菌细胞代谢活性(P<0.05),从而抑制生物膜形成;Jiang等[69]研究表明分离纯化自L. paracasei LS-6的细菌素LSX01可降低S. aureus浮游菌的活性,显著降低S. aureus生物膜密度(P<0.05);Li等[71]通过XTT还原试验结合荧光显微镜观察发现来自L. salivarius CGMCC2070的细菌素XJS01可使S. aureus细胞变形、表面形成孔洞并导致内容物流出,显著抑制S. aureus浮游菌和生物膜形成量(P<0.05);Xin等[93]研究表明纯化自L. plantarum LF-8的细菌素LFX01可使S. aureus浮游菌的死亡细胞数从3.04%显著增加至65.90%(P<0.05),代谢活力下降57.47%,从而降低生物膜密度;江宇航等[94]研究表明布氏乳杆菌(Lactobacillus buchneri)所产细菌素BSX01在1/2 MIC时可有效降低S. aureus生物膜的形成,在2 MIC时能够完全抑制生物膜的形成;Chen等[95]研究表明细菌素CLK_01(纯化自L. rhamnosus CLK101)可降低S. aureus浮游菌代谢活力,使浮游菌细胞皱缩、表面穿孔,并以浓度依赖性的方式降低S. aureus生物膜密度;许晓燕等[96]研究表明产自乳酸乳球菌乳亚种(Lactococcus lactis subsp. lactis)NCU036018的乳球菌素036018能使S. aureus生物膜形成率降至25.51%;杜贺超[97]研究了产自L. plantarum GZ1-27的plantaricin GZ1-27对耐甲氧西林金黄色葡萄球菌(Methicillin-resistant Staphylococcus aureus,MRSA)生物膜的抑制作用,结果表明1/2 MIC的plantaricin GZ1-27可显著抑制MRSA胞外聚合物的分泌(P<0.05),使生物膜形成量下降55.3%,还可通过杀死成熟生物膜内细菌而破坏生物膜。
5.2.2 乳酸菌细菌素对金黄色葡萄球菌生物膜合成的群体感应调控机制
金黄色葡萄球菌形成生物膜后会增加食品污染的风险,增强细菌的致病性[3]。其生物膜的合成涉及多个系统的调控,如QS系统、多糖细胞间黏附素(Polysaccharide intercellular adhesin,PIA)调控、胞外聚合物的合成等,其中QS系统对金黄色葡萄球菌生物膜合成的调控作用尤为重要。QS系统主要涉及两种调控因子,分别为辅助调节因子(Auxiliary gene regulation,Agr)和自体诱导物(Autoinducer-2,AI-2),这两种调控因子介导的群体感应系统以不同的方式调控生物膜的形成[98]。
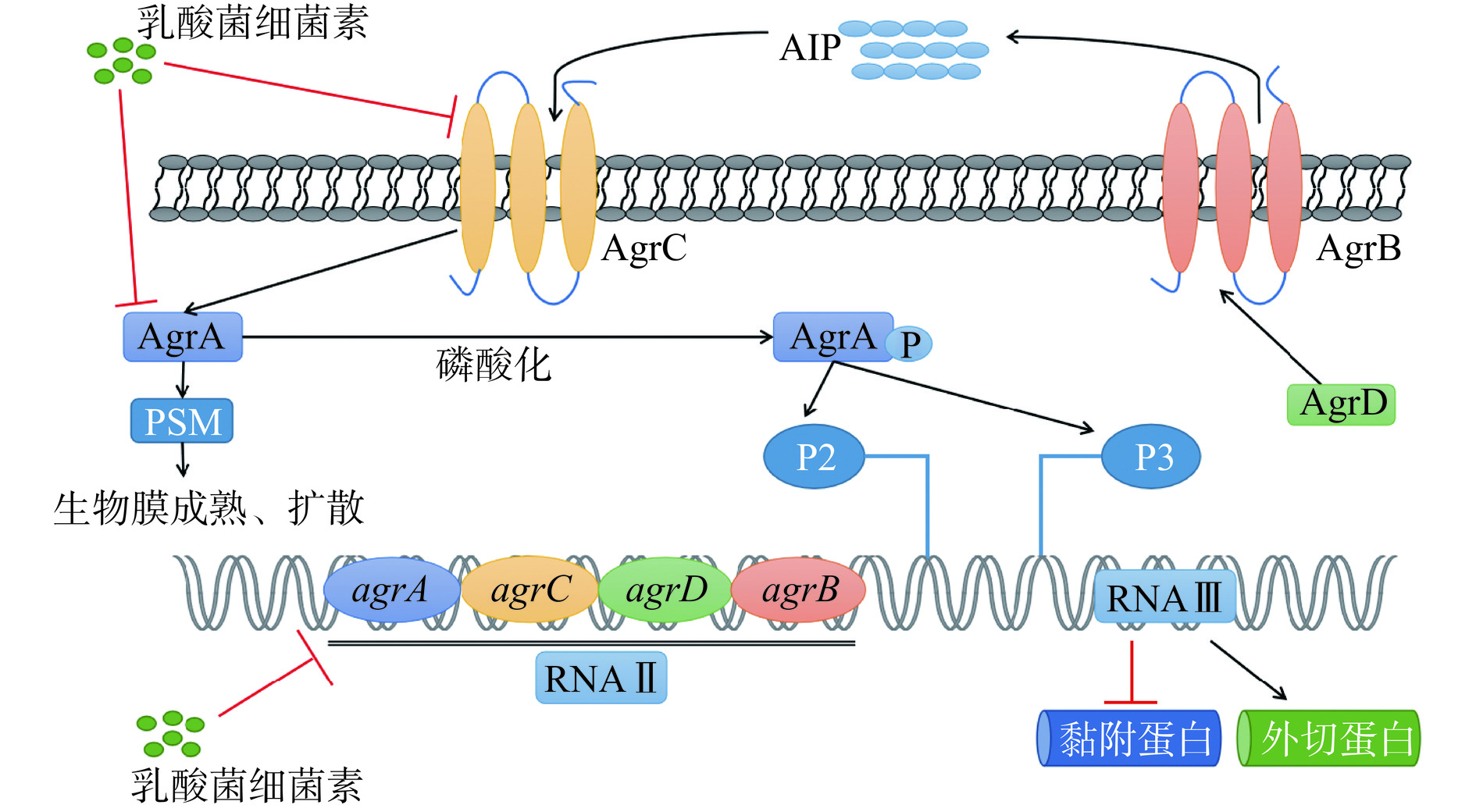
Agr系统主要调控细菌粘附和表面蛋白的表达,影响生物膜的粘附、成熟和分散过程[99]。Agr系统由RNAⅡ和RNAⅢ两个转录单位组成,P2和P3启动子分别驱动RNAII和RNAIII的转录。RNAII中含有4个部分,分别为AgrA、AgrB、AgrC和AgrD。AIP是Agr系统的信号分子,当AIP浓度达到一定阈值时,会与AgrC结合,进而使AgrA磷酸化并激活P2和P3启动子,启动agrA-D和RNAⅢ的转录。RNAIII是Agr系统的主要效应分子[100],上调编码外切蛋白(包括溶血素、毒素和外切蛋白酶)的基因,下调编码表面粘附素的基因[101−102]。表面活性剂酚溶性调节蛋白(Phenolsoluble modulins,PSMs)是S. aureus生物膜分散和传播的关键效应分子[103],AgrA与其启动子结合可产生PSMs[104−105],agr基因的上调会导致PSMs的增加,并促进生物膜的成熟和扩散。有研究表明乳酸菌细菌素可通过抑制S. aureus Agr系统中相关基因和相关蛋白的表达而抑制其生物膜的形成,例如Li等[106]研究表明新型乳源抗菌肽BCp12可与Agr系统的两种蛋白(AgrA和AgrC)结合并使生物膜形成相关基因agrA、agrB、agrC和psmβ的表达水平均下调,抑制S. aureus生物膜的形成。乳酸菌细菌素对金黄色葡萄球菌Agr系统的调控机制如图4所示。
![]() 图 4 乳酸菌细菌素对金黄色葡萄球菌Agr系统的影响Figure 4. Effect of lactic acid bacteria bacteriocins on the Agr system of Staphylococcus aureus
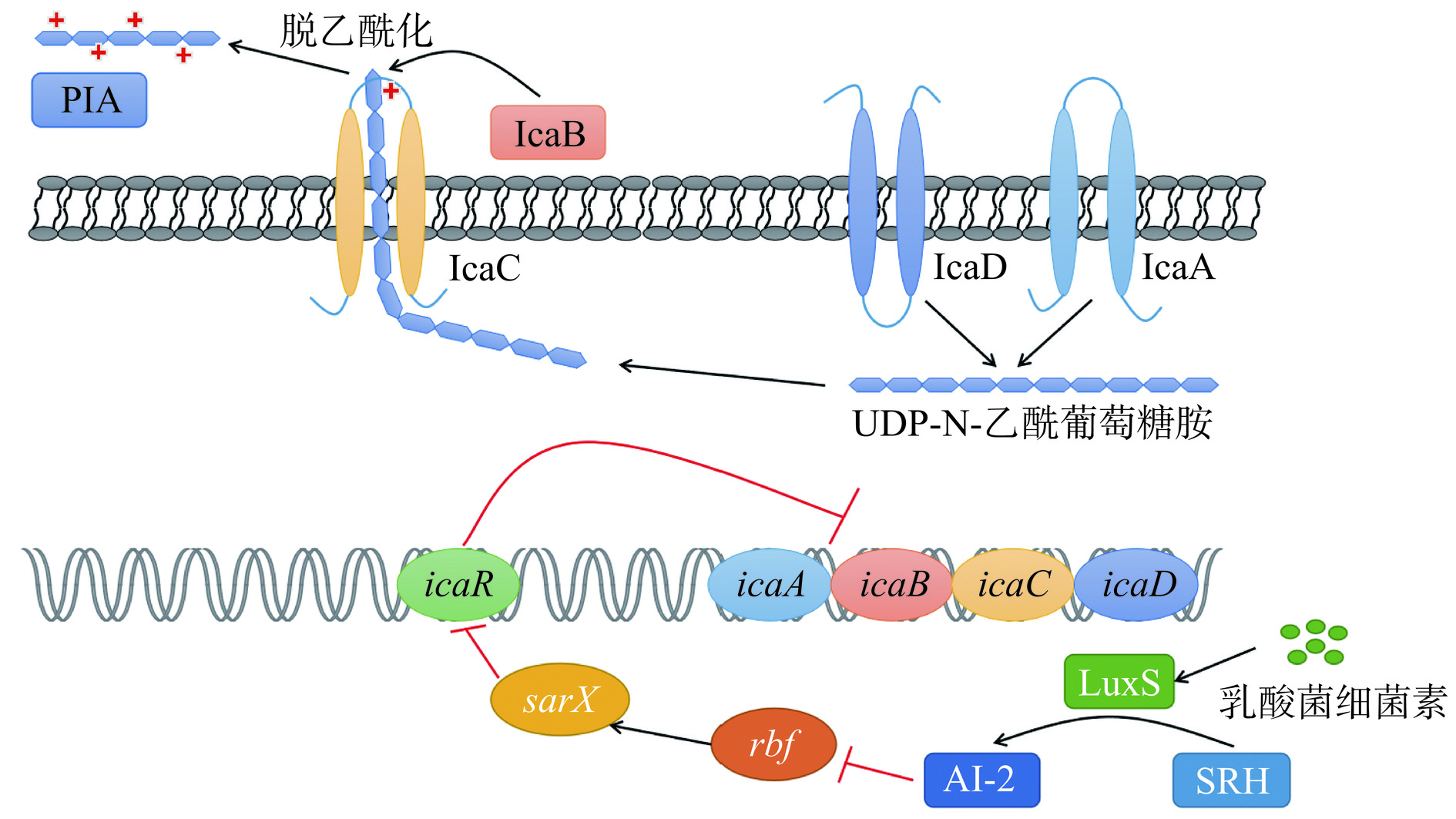
图 4 乳酸菌细菌素对金黄色葡萄球菌Agr系统的影响Figure 4. Effect of lactic acid bacteria bacteriocins on the Agr system of Staphylococcus aureusLuxS/AI-2系统主要通过调节多糖细胞间粘附素PIA合成来调控生物膜的形成。PIA是S. aureus生物膜胞外基质的重要成分,在定植、生物膜形成、抗生素耐药性等多个方面发挥重要作用[107]。PIA的合成由ica操纵子介导,ica操纵子由功能基因icaA、icaB、icaC、icaD和阻遏基因icaR共同构成[108]。icaR位于ica操纵子的上游,对PIA的合成基因icaA和icaD的表达具有抑制作用,可减少PIA的合成,抑制生物膜的形成[98]。LuxS/AI-2系统中LuxS可以催化SRH合成AI-2并抑制rbf基因的表达,从而促进sarX基因表达,激活icaR基因转录,减少生物膜形成[108]。有研究表明乳酸菌细菌素可使S. aureus的luxS基因表达增加从而抑制S. aureus生物膜形成,如L. plantarum YKX产生的plantaricin YKX能与AI-2分子及其合成酶LuxS结合,并使S. aureus 的luxS基因表达量显著增加(P<0.05),表明plantaricin YKX能通过调控S. aureus的LuxS/AI-2系统抑制其生物膜的形成[109];将产自L. rhamnosus XN2的细菌素XN2作用于S. aureus,发现1/2 MIC的细菌素XN2处理可使S. aureus中luxS基因表达量增加2倍,生物膜抑制率可达到45.8%±3.1%[110]。乳酸菌细菌素对金黄色葡萄球菌LuxS/AI-2系统的调控机制如图5所示。
![]() 图 5 乳酸菌细菌素对金黄色葡萄球菌LuxS/AI-2系统的影响Figure 5. Effect of lactic acid bacteria bacteriocins on the LuxS/AI-2 system of Staphylococcus aureus
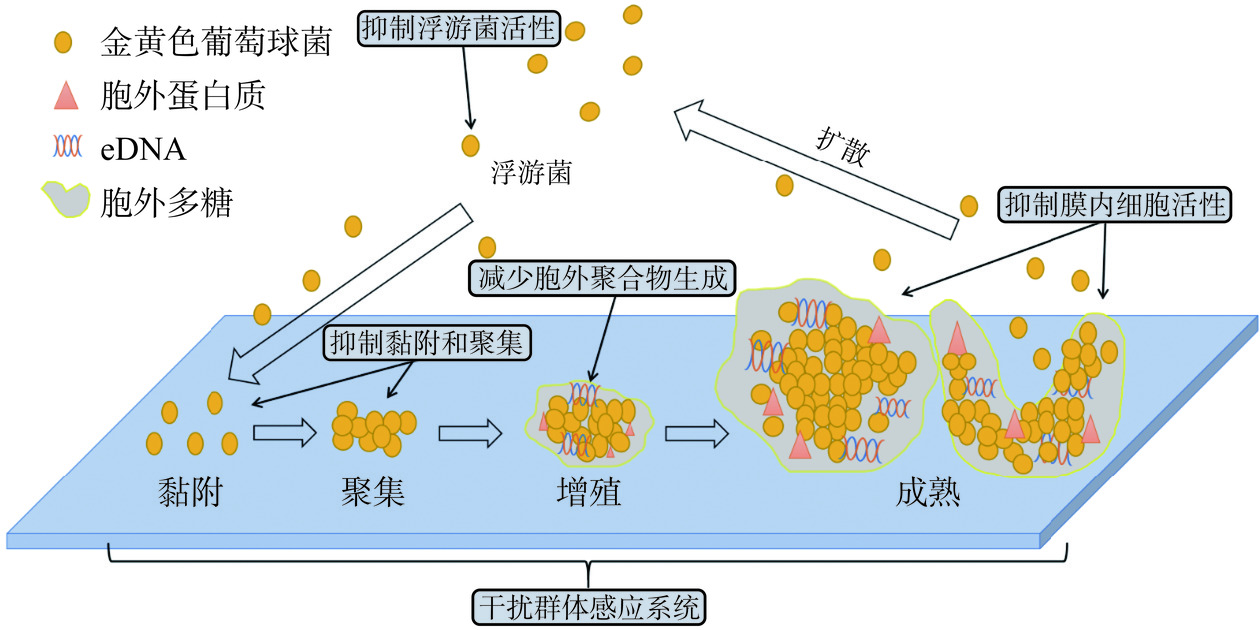
图 5 乳酸菌细菌素对金黄色葡萄球菌LuxS/AI-2系统的影响Figure 5. Effect of lactic acid bacteria bacteriocins on the LuxS/AI-2 system of Staphylococcus aureus综上可见,乳酸菌细菌素可通过杀死S. aureus浮游菌、降低黏附作用、抑制胞外聚合物的分泌、下调生物膜形成相关基因的表达及抑制膜内细胞活性从而减少生物膜的形成、破坏成熟生物膜;此外,乳酸菌细菌素也可通过调控S. aureus的QS系统而抑制其生物膜形成。据此结论得出乳酸菌细菌素对S. aureus生物膜的抑制机制如图6所示。
![]() 图 6 乳酸菌细菌素对金黄色葡萄球菌生物膜的抑制机制Figure 6. Mechanism of inhibition of Staphylococcus aureus biofilm by lactic acid bacteria bacteriocins
图 6 乳酸菌细菌素对金黄色葡萄球菌生物膜的抑制机制Figure 6. Mechanism of inhibition of Staphylococcus aureus biofilm by lactic acid bacteria bacteriocins6. 结论与展望
S. aureus及其生物膜的形成威胁着食品工业经济发展及食品安全,而传统处理方法易产生副作用,危害人体健康,因此,寻找有效、天然的抗生物膜策略十分必要。近年来许多研究表明乳酸菌细菌素有潜力作为天然防腐剂应用于食品,其抗菌和抗生物膜机制的研究也逐渐成为新的研究热点。目前大多乳酸菌细菌素对S. aureus生物膜抑制机制的研究仅停留在生物膜形成初期的浮游菌阶段,少数研究者分析了乳酸菌细菌素对S. aureus生物膜黏附、胞外聚合物分泌、成熟生物膜及生物膜形成相关基因和蛋白表达的影响。除了QS系统外,PIA和全局调控系统也可调控S. aureus生物膜的形成。因此,未来研究有必要从更多方面(如对PIA依赖性生物膜的影响、对全局调控系统的影响等方面)研究乳酸菌细菌素对S. aureus生物膜形成的调控机制。此外,结合基因组学、转录组学、蛋白质组学和代谢组学等工具或许有助于进一步研究乳酸菌细菌素的抗生物膜机制。明确乳酸菌细菌素的抗生物膜机制将有助于更多乳酸菌细菌素商品化,助力食品工业更好地控制S. aureus生物膜,为消费者提供品质更好、更加健康的食品。
-
![]()
图 2 群体感应调控下的细菌素合成与转运机制
Figure 2. Mechanism of bacteriocins synthesis and transport regulated by quorum sensing
![]()
图 4 乳酸菌细菌素对金黄色葡萄球菌Agr系统的影响
Figure 4. Effect of lactic acid bacteria bacteriocins on the Agr system of Staphylococcus aureus
![]()
图 5 乳酸菌细菌素对金黄色葡萄球菌LuxS/AI-2系统的影响
Figure 5. Effect of lactic acid bacteria bacteriocins on the LuxS/AI-2 system of Staphylococcus aureus
![]()
图 6 乳酸菌细菌素对金黄色葡萄球菌生物膜的抑制机制
Figure 6. Mechanism of inhibition of Staphylococcus aureus biofilm by lactic acid bacteria bacteriocins
表 2 细菌素粗提方法
Table 2 Crude extraction methods of bacteriocins
方法 原理 特点 文献 硫酸铵沉淀法 高浓度盐破坏蛋白质分子表面的水化层,与蛋白质竞争水分子,使蛋白质溶解度降低并形成沉淀,从而从溶液中
分离出来优点:溶解度高、成本低、操作简单、蛋白质在
高浓度盐条件下不易变性
缺点:粗提产物含大量杂蛋白,后期纯化难度大[55] 有机溶剂萃取法 基于细菌素的极性,将有机溶剂与细菌素混合,利用相似相溶原理将细菌素从水相溶解进入有机相,从而实现
细菌素的浓缩优点:有机溶剂易于除去
缺点:成本高、有机溶剂后处理时对环境的
污染问题[56−57] 细胞吸附/解吸法 利用细菌素的带电特性,调节溶液的pH可使菌体细胞吸附细菌素,再调回溶液原来的pH便可使细菌素解吸 优点:成本低和利于后期进一步纯化
缺点:有一定局限性,不具有普遍性[58] 膜分离技术 依据细菌素的分子量大小,选用合适的膜进行截留,达到分离的目的 优点:高选择性、不需要添加剂、易于扩大规模
缺点:样品损失量多,不易清洁[59]  下载: 导出CSV
下载: 导出CSV
表 3 细菌素纯化常规方法
Table 3 Bacteriocins purification conventional method
纯化方法 微生物菌株及来源 细菌素 分子量 文献 疏水相互作用色谱、固相萃取和RP-HPLC 发酵乳杆菌(Lactobacillus
Fermentum, L. Fermentum)GA715,羊乳Fermencin SA715 1792.537 Da [52] 凝胶过滤色谱和HPLC L. casei LiN333,江水菜 细菌素LiN333 4890 Da [66] 硫酸铵沉淀、pH介导的细胞吸附-解吸、阳离子交换色谱和RP-HPLC L. rhamnosus LS-8,四川泡菜 细菌素pH 25、S68、S81和S137 / [65] 硫酸铵沉淀、阳离子交换色谱和RP-HPLC L. plantarumSHY 21-2,牦牛酸奶 Plantaricin LP 21-2 1362.84 Da [67] 乙酸乙酯萃取、阴离子交换色谱和HPLC L. rhamnosus zrx01,Kefir 细菌素zrx01 / [68] 阳离子交换色谱和疏水相互作用色谱 L. plantarum GCNRC_GA15,埃及山羊奶酪 细菌素GA15 4369 Da [5] Superdex 30 Increase 10/300 GL排阻色谱 L. paracasei LS-6,发酵酸奶 细菌素LSX01 967.49 Da [69] 大孔树脂XAD-16吸附色谱、阳离子交换色谱、Sephadex G-25凝胶过滤色谱和RP-HPLC L. paracasei ZFM54,新生婴儿粪便 细菌素ZFM54 1143.39 Da [70] 乙酸乙酯萃取和阴离子交换色谱 L. rhamnosus 1.0320,酸马奶 细菌素1.0320 1~3.3 kDa [57] Superdex™ 30 Increase 10/300 GL排阻色谱 唾液乳杆菌(Lactobacillus salivarius, L. salivarius)CGMCC2070,云南黑骨鸡肠道粘膜 细菌素XJS01 666.31 Da [71] Ni-NTA亲和层析和RP-HPLC 结壳乳杆菌(Lactobacillus crustorum, L. crustorum)MN047,发酵马奶 细菌素BM173 / [72] 生物色谱法和RP-HPLC L. plantarum SLG10,康普茶 细菌素SLG10 1422 Da [64] 硫酸铵沉淀 L. plantarum PKLP5,生牛奶 细菌素Bac23 5.1 kDa [73] 乙酸乙酯萃取、Sephadex LH-20凝胶过滤色谱、阳离子交换色谱和RP-HPLC纯化 戊糖片球菌(Pediococcus pentosaceus)zy–B,
华贵栉孔扇贝肠道细菌素PE-ZYB1 2019.22 Da [56] 硫酸铵沉淀、阳离子交换色谱和RP-HPLC L. Fermentum BZ532,发酵谷物饮料 细菌素LF-BZ532 1105.563 Da [74] 双水相系统、Sephadex G - 50凝胶过滤色谱和RP-HPLC L. plantarum M1-UVs300,发酵香肠 细菌素M1-UVs300 3.4 kDa [75] 硫酸铵沉淀、固相萃取、阳离子交换色谱和RP - HPLC 屎肠球菌(Enterococcus faecium)HDX-2,腌渍黄瓜 肠道菌素HDX-2 5482 Da [76] 磁性脂质体吸附和RP-HPLC L. plantarum SLG1,牦牛干酪 Plantaricin SLG1 1083.25 Da [77] 硫酸铵沉淀和凝胶过滤色谱 L. lactis CH3,发酵乳制品 细菌素CH3 3.5 kDa [78]
下载: 导出CSV
-
[1] ZHENG J X, SHANG Y P, WU Y, et al. Loratadine inhibits Staphylococcus aureus virulence and biofilm formation[J]. iScience,2022,25(2):103731. doi: 10.1016/j.isci.2022.103731
[2] HOU W W, KANG S M, CHANG J, et al. Correlation analysis between GlpQ-Regulated degradation of wall teichoic acid and biofilm formation triggered by Lactobionic Acid in Staphylococcus aureus[J]. Foods,2022,11(21):3438. doi: 10.3390/foods11213438
[3] SHIN H J, YANG S, LIM Y. Antibiotic susceptibility of Staphylococcus aureus with different degrees of biofilm formation[J]. Journal of Analytical Science and Technology,2021,12(1):41. doi: 10.1186/s40543-021-00294-2
[4] COTTER P D, HILL C, ROSS R P. Bacteriocins:Developing innate immunity for food[J]. Nature Reviews Microbiology,2005,3(10):777−788. doi: 10.1038/nrmicro1273
[5] DABA G M, MOSTAFA F A, SALEH S A A, et al. Purification, amino acid sequence, and characterization of bacteriocin GA15, a novel class IIa bacteriocin secreted by Lactiplantibacillus plantarum GCNRC_GA15[J]. International Journal of Biological Macromolecules,2022,213:651−662. doi: 10.1016/j.ijbiomac.2022.06.003
[6] PANG X Y, SONG X Y, CHEN M J, et al. Combating biofilms of foodborne pathogens with bacteriocins by lactic acid bacteria in the food industry[J]. Comprehensive Reviews in Food Science and Food Safety,2022,21(2):1657−1676. doi: 10.1111/1541-4337.12922
[7] ZAPAŚNIK A, SOKOŁOWSKA B, BRYŁA M. Role of Lactic Acid Bacteria in food preservation and safety[J]. Foods,2022,11(9):1283. doi: 10.3390/foods11091283
[8] AHMAD V, KHAN M S, JAMAL Q M S, et al. Antimicrobial potential of bacteriocins:In therapy, agriculture and food preservation[J]. International Journal of Antimicrobial Agents,2017,49(1):1−11. doi: 10.1016/j.ijantimicag.2016.08.016
[9] EGAN K, FIELD D, REA M C, et al. Bacteriocins:novel solutions to age old spore-related problems?[J]. Frontiers in Microbiology, 2016, 7.
[10] MOKOENA M P. Lactic Acid Bacteria and their bacteriocins:classification, biosynthesis and applications against uropathogens:A mini-review[J]. Molecules,2017,22(8):1255. doi: 10.3390/molecules22081255
[11] PEREZ R H, ZENDO T, SONOMOTO K. Circular and leaderless bacteriocins:Biosynthesis, mode of action, applications, and prospects[J]. Frontiers in Microbiology,2018,9:2085. doi: 10.3389/fmicb.2018.02085
[12] JOHNSON E M, JUNG Dr Y G, JIN Dr Y Y, et al. Bacteriocins as food preservatives:Challenges and emerging horizons[J]. Critical Reviews in Food Science and Nutrition,2018,58(16):2743−2767. doi: 10.1080/10408398.2017.1340870
[13] ENRIQUETA G G, MAYER M J, COTTER P D, et al. Gut microbiota as a source of novel antimicrobials[J]. Gut Microbes,2019,10(1):1−21. doi: 10.1080/19490976.2018.1455790
[14] 李萍, 龙春昊, 赵轩, 等. Ⅱ类细菌素的生物合成及其在食品领域的应用[J]. 中国食品学报,2021,21(10):269−286. [LI P, LONG C H, ZHAO X, et al. Biosynthesis of class II bacteriocins and their applications in food[J]. Journal of Chinese Institute of Food Science and Technology,2021,21(10):269−286.] LI P, LONG C H, ZHAO X, et al. Biosynthesis of class II bacteriocins and their applications in food[J]. Journal of Chinese Institute of Food Science and Technology, 2021, 21(10): 269−286.
[15] YI Y L, LI P, ZHAO F, et al. Current status and potentiality of class II bacteriocins from lactic acid bacteria:Structure, mode of action and applications in the food industry[J]. Trends in Food Science & Technology,2022,120:387−401.
[16] ALVAREZ-SIEIRO P, MONTALBÁN-LÓPEZ M, MU D, et al. Bacteriocins of lactic acid bacteria:extending the family[J]. Applied Microbiology and Biotechnology,2016,100(7):2939−2951. doi: 10.1007/s00253-016-7343-9
[17] PÉREZ-RAMOS A, MADI-MOUSSA D, COUCHENEY F, et al. Current knowledge of the mode of action and immunity mechanisms of LAB-bacteriocins[J]. Microorganisms,2021,9(10):2107. doi: 10.3390/microorganisms9102107
[18] LI Y J, YU S, WENG P F, et al. Purification and antimicrobial mechanism of a novel bacteriocin produced by Lactiplantibacillus plantarum FB-2[J]. LWT,2023,185:115123. doi: 10.1016/j.lwt.2023.115123
[19] HAN X P, ZHANG M Y, PENG J Y, et al. Purification and characterization of a novel bacteriocin from Lactiplantibacillus plantarum Z057, and its antibacterial and antibiofilm activities against Vibrio parahaemolyticus[J]. LWT,2023,173:114358. doi: 10.1016/j.lwt.2022.114358
[20] LIU G R, NIE R, LIU Y S, et al. Combined antimicrobial effect of bacteriocins with other hurdles of physicochemic and microbiome to prolong shelf life of food:A review[J]. Science of The Total Environment,2022,825:154058. doi: 10.1016/j.scitotenv.2022.154058
[21] FU Y, ZHAO D B, WANG L Y, et al. A broad-spectrum novel bacteriocin produced by Lactobacillus sakei in Nanjing Steamed Roast Duck:Purification, antimicrobial characteristics, and antibacterial mechanisms[J]. Food Bioscience,2022,50:101995. doi: 10.1016/j.fbio.2022.101995
[22] YI L H, LUO L L, CHEN J X, et al. Cell wall and DNA damage of Staphylococcus aureus by bacteriocin BM1157[J]. LWT,2020,134:109842. doi: 10.1016/j.lwt.2020.109842
[23] RENDUELES C, DUARTE A C, ESCOBEDO S, et al. Combined use of bacteriocins and bacteriophages as food biopreservatives:A review[J]. International Journal of Food Microbiology,2022,368:109611. doi: 10.1016/j.ijfoodmicro.2022.109611
[24] JUTURU V, WU J C. Microbial production of bacteriocins:Latest research development and applications[J]. Biotechnology Advances,2018,36(8):2187−2200. doi: 10.1016/j.biotechadv.2018.10.007
[25] VERMA D K, THAKUR M, SINGH S, et al. Bacteriocins as antimicrobial and preservative agents in food:Biosynthesis, separation and application[J]. Food Bioscience,2022,46:101594. doi: 10.1016/j.fbio.2022.101594
[26] KIRTONIA K, SALAUDDIN M, BHARADWAJ K K, et al. Bacteriocin:A new strategic antibiofilm agent in food industries[J]. Biocatalysis and Agricultural Biotechnology,2021,36:102141. doi: 10.1016/j.bcab.2021.102141
[27] ZHU L Y, ZENG J W, WANG C, et al. structural basis of pore formation in the mannose phosphotransferase system by pediocin PA-1[J]. Applied and Environmental Microbiology,2022,88(3):e01992−21.
[28] CHELLIAH R, SARAVANAKUMAR K, DALIRI E B M, et al. Unveiling the potentials of bacteriocin (Pediocin L50) from Pediococcus acidilactici with antagonist spectrum in a Caenorhabditis elegans model[J]. International Journal of Biological Macromolecules,2020,143:555−572. doi: 10.1016/j.ijbiomac.2019.10.196
[29] SILPA S, RUPACHANDRA S. Cyclic peptide production from lactic acid bacteria (LAB) and their diverse applications[J]. Critical Reviews in Food Science and Nutrition,2022,62(11):2909−2927. doi: 10.1080/10408398.2020.1860900
[30] KAREB O, AÏDER M. Quorum Sensing circuits in the communicating mechanisms of bacteria and its implication in the biosynthesis of bacteriocins by lactic acid bacteria:A review[J]. Probiotics and Antimicrobial Proteins,2020,12(1):5−17. doi: 10.1007/s12602-019-09555-4
[31] MILLER E L, KJOS M, ABRUDAN M I, et al. Eavesdropping and crosstalk between secreted quorum sensing peptide signals that regulate bacteriocin production in Streptococcus pneumoniae[J]. The ISME Journal,2018,12(10):2363−2375. doi: 10.1038/s41396-018-0178-x
[32] GARCÍA-CURIEL L, DEL ROCÍO LÓPEZ-CUELLAR Ma, RODRÍGUEZ-HERNÁNDEZ A I, et al. Toward understanding the signals of bacteriocin production by Streptococcus spp. and their importance in current applications[J]. World Journal of Microbiology and Biotechnology,2021,37(1):15. doi: 10.1007/s11274-020-02973-5
[33] TURAN N B, CHORMEY D S, BUYUKPINAR C, et al. Quorum sensing:Little talks for an effective bacterial coordination[J]. Trac-Trends in Analytical Chemistry,2017,91:1−11. doi: 10.1016/j.trac.2017.03.007
[34] FREEMAN J A, BASSLER B L. Sequence and function of LuxU:A two-component phosphorelay protein that regulates quorum sensing in Vibrio harveyi.[J]. Journal of Bacteriology,1999,181(3):899−906. doi: 10.1128/JB.181.3.899-906.1999
[35] MALDONADO-BARRAGÁN A, CABALLERO-GUERRERO B, LUCENA-PADRÓS H, et al. Induction of bacteriocin production by coculture is widespread among plantaricin-producing Lactobacillus plantarum strains with different regulatory operons[J]. Food Microbiology,2013,33(1):40−47. doi: 10.1016/j.fm.2012.08.009
[36] QIAN X, TIAN P J, ZHAO J X, et al. Quorum sensing of lactic acid bacteria:Progress and insights[J]. Food Reviews International,2023,39(7):4781−4792. doi: 10.1080/87559129.2022.2062766
[37] AXELSSON L, HOLCK A. The genes involved in production of and immunity to sakacin A, a bacteriocin from Lactobacillus sake Lb706[J]. Journal of Bacteriology,1995,177(8):2125−2137. doi: 10.1128/jb.177.8.2125-2137.1995
[38] ISHIBASHI N, MATSUMOTO N, PEREZ R H, et al. Molecular characterization of the possible regulation of multiple bacteriocin production through a three-component regulatory system in Enterococcus faecium NKR-5-3[J]. Journal of Bioscience and Bioengineering,2021,131(2):131−138. doi: 10.1016/j.jbiosc.2020.09.019
[39] DE KEERSMAECKER S C J, VARSZEGI C, VAN BOXEL N, et al. Chemical synthesis of (S)-4, 5-dihydroxy-2, 3-pentanedione, a bacterial signal molecule precursor, and validation of its activity in Salmonella typhimurium[J]. Journal of Biological Chemistry,2005,280(20):19563−19568. doi: 10.1074/jbc.M412660200
[40] MENG F Q, ZHAO M W, LU Z X. The LuxS/AI-2 system regulates the probiotic activities of lactic acid bacteria[J]. Trends in Food Science & Technology,2022,127:272−279.
[41] JOHANSEN P, JESPERSEN L. Impact of quorum sensing on the quality of fermented foods[J]. Current Opinion in Food Science,2017,13:16−25. doi: 10.1016/j.cofs.2017.01.001
[42] JIA F F, PANG X H, ZHU D Q, et al. Role of the luxS gene in bacteriocin biosynthesis by Lactobacillus plantarum KLDS1.0391:A proteomic analysis[J]. Scientific Reports,2017,7(1):13871. doi: 10.1038/s41598-017-13231-4
[43] LIU G R, NIE R, LIU Y S, et al. Bacillus subtilis BS-15 effectively improves plantaricin production and the regulatory biosynthesis in Lactiplantibacillus plantarum RX-8[J]. Frontiers in Microbiology,2022,12:772546. doi: 10.3389/fmicb.2021.772546
[44] MAN L L, XIANG D J. LuxS-mediated quorum sensing system in Lactobacillus plantarum NMD-17 from koumiss:Induction of plantaricin MX in co-cultivation with certain lactic acid bacteria[J]. Folia Microbiologica,2021,66(5):855−871.
[45] LI J P, YANG X Y, SHI G C, et al. Cooperation of lactic acid bacteria regulated by the AI-2/LuxS system involve in the biopreservation of refrigerated shrimp[J]. Food Research International,2019,120:679−687. doi: 10.1016/j.foodres.2018.11.025
[46] GARSA A K, KUMARIYA R, SOOD S K, et al. Bacteriocin production and different strategies for their recovery and purification[J]. Probiotics and Antimicrobial Proteins,2014,6(1):47−58. doi: 10.1007/s12602-013-9153-z
[47] ALZBUTAS G, KANIUSAITE M, LAGUNAVICIUS A. Enhancement of DNaseI salt tolerance by mimicking the domain structure of dnase from an extremely halotolerant bacterium Thioalkalivibrio sp. K90mix[J]. PLOS ONE,2016,11(3):e0150404. doi: 10.1371/journal.pone.0150404
[48] 黄倩, 万倩, 刘爽, 等. 抑制酸奶后酸化产细菌素乳酸菌的筛选及其产细菌素条件优化[J]. 西南民族大学学报(自然科学版),2023,49(5):482−492. [HUANG Q, WAN Q, LIU S, et al. Screening of bacteriocin-producing lactic acid bacteria to inhibit post-acidification of yogurt and optimization of bacteriocin-producing conditions[J]. Journal of Southwest University for Nationalities (Natural Science Edition),2023,49(5):482−492.] doi: 10.11920/xnmdzk.2023.05.002 HUANG Q, WAN Q, LIU S, et al. Screening of bacteriocin-producing lactic acid bacteria to inhibit post-acidification of yogurt and optimization of bacteriocin-producing conditions[J]. Journal of Southwest University for Nationalities (Natural Science Edition), 2023, 49(5): 482−492. doi: 10.11920/xnmdzk.2023.05.002
[49] 唐美. 卷曲乳杆菌发酵培养基优化及其抑菌物质的初步分离纯化和鉴定[D]. 长沙:中南大学, 2022. [TANG M. Optimization of fermentation medium of Lactobacillus curvatus and preliminary isolation, purification and identification of bacteriostatic substances[D]. Changsha:Central South University, 2022.] TANG M. Optimization of fermentation medium of Lactobacillus curvatus and preliminary isolation, purification and identification of bacteriostatic substances[D]. Changsha: Central South University, 2022.
[50] JAWAN R, ABBASILIASI S, TAN J S, et al. Influence of culture conditions and medium compositions on the production of bacteriocin-like inhibitory substances by Lactococcus lactis Gh1[J]. Microorganisms,2020,8(10):1454. doi: 10.3390/microorganisms8101454
[51] YE Z M, ZHOU X H, JIANG B T, et al. An optimal carbon source can enhance the Paracin 1.7 titer and reduce carbon source consumption in cocultures of Lacticaseibacillus paracasei HD1.7 and Bacillus spp.[J]. LWT,2022,161:113336. doi: 10.1016/j.lwt.2022.113336
[52] WAYAH S B, PHILIP K. Characterization, yield optimization, scale up and biopreservative potential of fermencin SA715, a novel bacteriocin from Lactobacillus fermentum GA715 of goat milk origin[J]. Microbial Cell Factories,2018,17(1):125. doi: 10.1186/s12934-018-0972-1
[53] PARLINDUNGAN E, DEKIWADIA C, JONES O A H. Factors that influence growth and bacteriocin production in Lactiplantibacillus plantarum B21[J]. Process Biochemistry,2021,107:18−26. doi: 10.1016/j.procbio.2021.05.009
[54] LI C, BAI J H, CAI Z L, et al. Optimization of a cultural medium for bacteriocin production by Lactococcus lactis using response surface methodology[J]. Journal of Biotechnology,2002,93(1):27−34. doi: 10.1016/S0168-1656(01)00377-7
[55] ZHANG J M, YANG Y Y, YANG H, et al. Purification and partial characterization of bacteriocin lac-b23, a novel bacteriocin production by Lactobacillus plantarum J23, isolated from chinese traditional fermented milk[J]. Frontiers in Microbiology,2018,9:2165. doi: 10.3389/fmicb.2018.02165
[56] ZHANG Y, YANG J M, LIU Y, et al. A novel bacteriocin PE-ZYB1 produced by Pediococcus pentosaceus zy-B isolated from intestine of Mimachlamys nobilis:Purification, identification and its anti-listerial action[J]. LWT,2020,118:108760. doi: 10.1016/j.lwt.2019.108760
[57] XU C, FU Y Y, LIU F, et al. Purification and antimicrobial mechanism of a novel bacteriocin produced by Lactobacillus rhamnosus 1.0320[J]. LWT,2021,137:110338. doi: 10.1016/j.lwt.2020.110338
[58] SADISHKUMAR V, JEEVARATNAM K. Purification and partial characterization of antilisterial bacteriocin produced by Pediococcus pentosaceus KJBC11 from Idli batter fermented with Piper betle leaves[J]. Journal of Food Biochemistry,2018,42(1):e12460. doi: 10.1111/jfbc.12460
[59] MURIANA P M, KLAENHAMMER T R. Purification and partial characterization of lactacin F, a bacteriocin produced by Lactobacillus acidophilus 11088[J]. Applied and Environmental Microbiology,1991,57(1):114−121. doi: 10.1128/aem.57.1.114-121.1991
[60] 胡彦新. 食品乳杆菌FM-MM4产细菌素的分离纯化及其特性研究[D]. 南京:南京农业大学, 2019. [HU Y X. Isolation and purification of bacteriocin-producing Lactobacillus casei FM-MM4 and its characterization [D]. Nanjing:Nanjing Agricultural University, 2019.] HU Y X. Isolation and purification of bacteriocin-producing Lactobacillus casei FM-MM4 and its characterization [D]. Nanjing: Nanjing Agricultural University, 2019.
[61] 陆泉, 施波, 李瑞胜, 等. 离子交换色谱在细菌素分离纯化中的应用[J]. 中国微生态学杂志,2010,22(6):570−572. [LU Q, SHI B, LI R S, et al. Application of ion-exchange chromatography in the separation and purification of bacteriocins[J]. Chinese Journal of Microecology,2010,22(6):570−572.] LU Q, SHI B, LI R S, et al. Application of ion-exchange chromatography in the separation and purification of bacteriocins[J]. Chinese Journal of Microecology, 2010, 22(6): 570−572.
[62] ZOU J, JIANG H, CHENG H, et al. Strategies for screening, purification and characterization of bacteriocins[J]. International Journal of Biological Macromolecules,2018,117:781−789. doi: 10.1016/j.ijbiomac.2018.05.233
[63] JAMALUDDIN N, STUCKEY D C, ARIFF A B, et al. Novel approaches to purifying bacteriocin:A review[J]. Critical Reviews in Food Science and Nutrition,2018,58(14):2453−2465. doi: 10.1080/10408398.2017.1328658
[64] PEI J, JIN W, ABD EL-ATY A M, et al. Isolation, purification, and structural identification of a new bacteriocin made by Lactobacillus plantarum found in conventional kombucha[J]. Food Control,2020,110:106923. doi: 10.1016/j.foodcont.2019.106923
[65] GUO X, CHEN J X, SUN H M, et al. Mining, heterologous expression, purification and characterization of 14 novel bacteriocins from Lactobacillus rhamnosus LS-8[J]. International Journal of Biological Macromolecules,2020,164:2162−2176. doi: 10.1016/j.ijbiomac.2020.08.067
[66] ULLAH N, WANG X J, LI T, et al. Purification and primary characterization of a novel bacteriocin, LiN333, from Lactobacillus casei, an isolate from a Chinese fermented food[J]. LWT,2017,84:867−875. doi: 10.1016/j.lwt.2017.04.056
[67] PENG S D, SONG J J, ZENG W Y, et al. A broad-spectrum novel bacteriocin produced by Lactobacillus plantarum SHY 21–2 from yak yogurt:Purification, antimicrobial characteristics and antibacterial mechanism[J]. LWT,2021,142:110955. doi: 10.1016/j.lwt.2021.110955
[68] ZHAO R X, LU Y Q, RAN J J, et al. Purification and characterization of bacteriocin produced by Lactobacillus rhamnosus zrx01[J]. Food Bioscience,2020,38:100754. doi: 10.1016/j.fbio.2020.100754
[69] JIANG Y H, XIN W G, YANG L Y, et al. A novel bacteriocin against Staphylococcus aureus from Lactobacillus paracasei isolated from Yunnan traditional fermented yogurt:Purification, antibacterial characterization, and antibiofilm activity[J]. Journal of Dairy Science,2022,105(3):2094−2107. doi: 10.3168/jds.2021-21126
[70] ZHU Y Y, ZHOU Q Q, LI P, et al. Purification, characterization, and mode of action of Paracin 54, a novel bacteriocin against Staphylococci[J]. Applied Microbiology and Biotechnology,2021,105(18):6735−6748. doi: 10.1007/s00253-021-11505-6
[71] LI H W, XIANG Y Z, ZHANG M, et al. A novel bacteriocin from Lactobacillus salivarius against Staphylococcus aureus:Isolation, purification, identification, antibacterial and antibiofilm activity[J]. LWT,2021,140:110826. doi: 10.1016/j.lwt.2020.110826
[72] QIAO Z, CHEN J X, ZHOU Q Q, et al. Purification, characterization, and mode of action of a novel bacteriocin BM173 from Lactobacillus crustorum MN047 and its effect on biofilm formation of Escherichia coli and Staphylococcus aureus[J]. Journal of Dairy Science,2021,104(2):1474−1483. doi: 10.3168/jds.2020-18959
[73] SIDHU P K, NEHRA K. Purification and characterization of bacteriocin Bac23 extracted from Lactobacillus plantarum PKLP5 and its interaction with silver nanoparticles for enhanced antimicrobial spectrum against food-borne pathogens[J]. LWT,2021,139:110546. doi: 10.1016/j.lwt.2020.110546
[74] RASHEED H A, TUOHETI T, ZHANG Y, et al. Purification and partial characterization of a novel bacteriocin produced by bacteriocinogenic Lactobacillus fermentum BZ532 isolated from Chinese fermented cereal beverage (Bozai)[J]. LWT,2020,124:109113. doi: 10.1016/j.lwt.2020.109113
[75] AN Y, WANG Y, LIANG X Y, et al. Purification and partial characterization of M1-UVs300, a novel bacteriocin produced by Lactobacillus plantarum isolated from fermented sausage[J]. Food Control,2017,81:211−217. doi: 10.1016/j.foodcont.2017.05.030
[76] DU R P, PING W X, GE J P. Purification, characterization and mechanism of action of enterocin HDX-2, a novel class IIa bacteriocin produced by Enterococcus faecium HDX-2[J]. LWT,2022,153:112451. doi: 10.1016/j.lwt.2021.112451
[77] PEI J, LI X S, HAN H, et al. Purification and characterization of plantaricin SLG1, a novel bacteriocin produced by Lb. plantarum isolated from yak cheese[J]. Food Control,2018,84:111−117. doi: 10.1016/j.foodcont.2017.07.034
[78] KRISHNAMOORTHI R, SRINIVASH M, MAHALINGAM P U, et al. Antimicrobial, anti-biofilm, antioxidant and cytotoxic effects of bacteriocin by Lactococcus lactis strain CH3 isolated from fermented dairy products—An in vitro and in silico approach[J]. International Journal of Biological Macromolecules,2022,220:291−306. doi: 10.1016/j.ijbiomac.2022.08.087
[79] ISABEL PEREZ-LOPEZ M, MENDEZ-REINA R, TRIER S, et al. Variations in carotenoid content and acyl chain composition in exponential, stationary and biofilm states of Staphylococcus aureus, and their influence on membrane biophysical properties[J]. Biochimica Et Biophysica ACTA-Biomembranes,2019,1861(5):978−987. doi: 10.1016/j.bbamem.2019.02.001
[80] GALIÉ S, GARCÍA-GUTIÉRREZ C, MIGUÉLEZ E M, et al. Biofilms in the food industry:Health aspects and control methods[J]. Frontiers in Microbiology,2018,9:898. doi: 10.3389/fmicb.2018.00898
[81] ABDALLAH M, BENOLIEL C, DRIDER D, et al. Biofilm formation and persistence on abiotic surfaces in the context of food and medical environments[J]. Archives of Microbiology,2014,196(7):453−472. doi: 10.1007/s00203-014-0983-1
[82] TITOUCHE Y, HAKEM A, HOUALI K, et al. Emergence of methicillin-resistant Staphylococcus aureus (MRSA) ST8 in raw milk and traditional dairy products in the Tizi Ouzou area of Algeria[J]. Journal of Dairy Science,2019,102(8):6876−6884. doi: 10.3168/jds.2018-16208
[83] PALÁ T R, SEVILLA A. Microbial contamination of carcasses, meat, and equipment from an Iberian pork cutting plant[J]. Journal of Food Protection,2004,67(8):1624−1629. doi: 10.4315/0362-028X-67.8.1624
[84] SHI X M, ZHU X N. Biofilm formation and food safety in food industries[J]. Trends in Food Science & Technology,2009,20(9):407−413.
[85] WANG H, HOU X, SHEN J W, et al. Alternative sigma factor B reduces biofilm formation and stress response in milk-derived Staphylococcus aureus[J]. LWT,2022,162:113515. doi: 10.1016/j.lwt.2022.113515
[86] WANG H W, WANG H H, LIANG L J, et al. Prevalence, genetic characterization and biofilm formation in vitro of Staphylococcus aureus isolated from raw chicken meat at retail level in Nanjing, China[J]. Food Control,2018,86:11−18. doi: 10.1016/j.foodcont.2017.10.028
[87] BESHIRU A, IGBINOSA I H, IGBINOSA E O. Characterization of enterotoxigenic Staphylococcus aureus from ready-to-eat seafood (RTES)[J]. LWT-Food Science and Technology,2021,135:110042. doi: 10.1016/j.lwt.2020.110042
[88] AL-SHABIB N A, HUSAIN F M, AHMAD I, et al. Eugenol inhibits quorum sensing and biofilm of toxigenic MRSA strains isolated from food handlers employed in Saudi Arabia[J]. BIOTECHNOLOGY & BIOTECHNOLOGICAL EQUIPMENT,2017,31(2):387−396.
[89] MIAO J, LIANG Y R, CHEN L Q, et al. Formation and development of Staphylococcus biofilm:With focus on food safety[J]. Journal of Food Safety,2017,37(4):e12358. doi: 10.1111/jfs.12358
[90] SUDAGIDAN M, YEMENICIOGLU A. Effects of Nisin and Lysozyme on growth inhibition and biofilm formation capacity of Staphylococcus aureus strains isolated from raw milk and cheese samples[J]. Journal of Food Protection,2012,75(9):1627−1633. doi: 10.4315/0362-028x.jfp-12-001
[91] ZHAO D Y, WANG Q, LU F X, et al. A novel plantaricin 827 effectively inhibits Staphylococcus aureus and extends shelf life of skim milk[J]. LWT,2022,154:112849. doi: 10.1016/j.lwt.2021.112849
[92] KIM J H, AHN H, LEE D, et al. Antibiofilm activity of crude bacteriocin JM01 produced by Pediococcus acidilactici against methicillin-resistant Staphylococcus aureus (MRSA)[J]. International Journal of Food Science and Technology,2023,58(5):2580−2589. doi: 10.1111/ijfs.16407
[93] XIN W G, WU G, YING J P, et al. Antibacterial activity and mechanism of action of bacteriocin LFX01 against Staphylococcus aureus and Escherichia coli and its application on pork model[J]. Meat Science,2023,196:109045. doi: 10.1016/j.meatsci.2022.109045
[94] 江宇航, 周寰宇, 辛维岗, 等. 布氏乳杆菌细菌素BSX01对金黄色葡萄球菌及其生物被膜形成的影响[J]. 食品与生物技术学报,2023,42(11):19−26. [JIANG Y H, ZHOU H Y, XIN W G, et al. Effects of Lactobacillus buchneri bacteriocin BSX01 on Staphylococcus aureus and its biofilm formation[J]. Journal of Food and Biotechnology,2023,42(11):19−26.] doi: 10.12441/spyswjs.20210419004 JIANG Y H, ZHOU H Y, XIN W G, et al. Effects of Lactobacillus buchneri bacteriocin BSX01 on Staphylococcus aureus and its biofilm formation[J]. Journal of Food and Biotechnology, 2023, 42(11): 19−26. doi: 10.12441/spyswjs.20210419004
[95] CHEN S Y, YANG R S, CI B Q, et al. A novel bacteriocin against multiple foodborne pathogens from Lacticaseibacillus rhamnosus isolated from juice ferments:ATF perfusion-based preparation of viable cells, characterization, antibacterial and antibiofilm activity[J]. Current Research in Food Science,2023,6:100484. doi: 10.1016/j.crfs.2023.100484
[96] 许晓燕, 彭珍, 熊世进, 等. 乳酸乳球菌乳亚种NCU036018细菌素的分离纯化及其抗菌机制[J]. 食品科学,2022,43(16):209−216. [XU X Y, PENG Z, XIONG S J, et al. Isolation and purification of bacteriocins from Lactococcus lactis subspecies NCU036018 and its antibacterial mechanism[J]. Food Science,2022,43(16):209−216.] doi: 10.7506/spkx1002-6630-20210902-018 XU X Y, PENG Z, XIONG S J, et al. Isolation and purification of bacteriocins from Lactococcus lactis subspecies NCU036018 and its antibacterial mechanism[J]. Food Science, 2022, 43(16): 209−216. doi: 10.7506/spkx1002-6630-20210902-018
[97] 杜贺超. Plantaricin GZ1-27的纯化鉴定及其对耐甲氧西林金黄色葡萄球菌抑菌机理的研究[D]. 南京:南京农业大学, 2022. [DU H C. Purification and characterization of Plantaricin GZ1-27 and its inhibitory mechanism against methicillin-resistant Staphylococcus aureus[D]. Nanjing:Nanjing Agricultural University, 2022.] DU H C. Purification and characterization of Plantaricin GZ1-27 and its inhibitory mechanism against methicillin-resistant Staphylococcus aureus[D]. Nanjing: Nanjing Agricultural University, 2022.
[98] LIU M H, WU X X, LI J K, et al. The specific anti-biofilm effect of gallic acid on Staphylococcus aureus by regulating the expression of the ica operon[J]. Food Control,2017,73:613−618. doi: 10.1016/j.foodcont.2016.09.015
[99] OTTO M. Staphylococcal infections:Mechanisms of biofilm maturation and detachment as critical determinants of pathogenicity[J]. Annual Review of Medicine,2013,64(1):175−188. doi: 10.1146/annurev-med-042711-140023
[100] JI G, BEAVIS R, NOVICK R P. Bacterial interference caused by autoinducing peptide variants[J]. Science,1997,276(5321):2027−2030. doi: 10.1126/science.276.5321.2027
[101] NOVICK R P, ROSS H F, PROJAN S J, et al. Synthesis of Staphylococcal virulence factors is controlled by a regulatory RNA molecule.[J]. The EMBO Journal,1993,12(10):3967−3975. doi: 10.1002/j.1460-2075.1993.tb06074.x
[102] CHEUNG G Y C, WANG R, KHAN B A, et al. Role of the accessory gene regulator agr in community-associated methicillin-resistant Staphylococcus aureus pathogenesis[J]. Infection and Immunity,2011,79(5):1927−1935. doi: 10.1128/IAI.00046-11
[103] PERIASAMY S, JOO H S, DUONG A C, et al. How Staphylococcus aureus biofilms develop their characteristic structure[J]. Proceedings of the National Academy of Sciences,2012,109(4):1281−1286.
[104] QUECK S Y, JAMESON-LEE M, VILLARUZ A E, et al. RNAIII-Independent target gene control by the agr Quorum-Sensing system:Insight into the evolution of virulence regulation in Staphylococcus aureus[J]. Molecular Cell,2008,32(1):150−158. doi: 10.1016/j.molcel.2008.08.005
[105] LE K Y, VILLARUZ A E, ZHENG Y, et al. Role of phenol-soluble modulins in staphylococcus epidermidis biofilm formation and infection of indwelling medical devices[J]. Journal of Molecular Biology,2019,431(16):3015−3027.
[106] LI Y F, LI S J, YANG K, et al. Antibiofilm mechanism of a novel milk-derived antimicrobial peptide against Staphylococcus aureus by downregulating agr quorum sensing system[J]. Journal of Applied Microbiology,2022,133(4):2198−2209.
[107] CRAMTON S E, ULRICH M, GÖTZ F, et al. anaerobic conditions induce expression of polysaccharide intercellular adhesin in Staphylococcus aureus and Staphylococcus epidermidis[J]. Infection and Immunity,2001,69(6):4079−4085. doi: 10.1128/IAI.69.6.4079-4085.2001
[108] CRAMTON S E, GERKE C, SCHNELL N F, et al. The intercellular adhesion (ica) locus is present in Staphylococcus aureus and is required for biofilm formation[J]. Infection and Immunity,1999,67(10):5427−5433. doi: 10.1128/IAI.67.10.5427-5433.1999
[109] PEI J J, HUANG Y G, REN T, et al. The antibacterial activity mode of action of plantaricin ykx against Staphylococcus aureus[J]. Molecules,2022,27(13):4280. doi: 10.3390/molecules27134280
[110] WEI Y H, WANG J Z, LIU Z, et al. Isolation and characterization of bacteriocin-producing Lacticaseibacillus rhamnosus XN2 from yak yoghurt and its bacteriocin[J]. Molecules,2022,27(7):2066. doi: 10.3390/molecules27072066
 下载:
下载:






计量
- 文章访问数: 0
- HTML全文浏览量: 0
- PDF下载量: 0
