


In Vitro Self-assembly Study of ZEN/DON Anti-idiotype Nanobody Based on SpyCatcher/SpyTag
-
摘要: 为了提升真菌毒素抗独特型纳米抗体的免疫分析性能,并实现双特异性检测抗原的模拟,本研究以玉米赤霉烯酮(Zearalenone,ZEN)、脱氧雪腐镰刀菌烯醇(Deoxynivalenol,DON)的抗独特型纳米抗体N28、Z6为研究对象,将SpyCatcher/SpyTag标签与N28、Z6分别进行融合表达,在此基础上基于体外自组装的方式,开展N28、Z6的双价、双特异性抗独特型纳米抗体的构建研究。结果表明,实现了四种SpyCatcher/SpyTag标签融合抗独特型纳米抗体N28-SpyCatcher、N28-SpyTag、Z6-SpyCatcher、Z6-SpyTag的原核可溶表达。基于Z6-SpyCatcher、Z6-SpyTag所建立的竞争抑制标准曲线的IC50分别为0.18、0.20 ng/mL,其灵敏度高于单体Z6的IC50(0.28 ng/mL);SpyCatcher标签与N28融合后有利于N28检测灵敏度的提升,IC50值由94.39降低至42.33 ng/mL,而SpyTag标签与N28融合后,则抑制了其与DON抗体的结合。此外,在4 ℃反应12 h的简单反应条件下,基于SpyCatcher/SpyTag的体外自组装特性,成功制备了Z6、N28的双价、双特异性抗独特型纳米抗体Z6-Z6、N28-28与N28-Z6、Z6-N28。ELISA的鉴定结果表明,双价抗独特型纳米抗体均保留了与对应抗体的结合特性,其中N28-Z6还实现了同时与ZEN、DON抗体的结合活性,由于SpyTag标签与N28结合会阻碍其与DON抗体的结合,因此Z6-N28仅保留了与ZEN抗体结合的活性。综上,本研究基于SpyCatcher/SpyTag实现了ZEN/DON双价、双特异性抗独特型纳米抗体的简便快速制备,同时提示SpyCatcher标签对于ZEN/DON抗独特型纳米抗体的免疫分析性能提升具有促进作用。
-
关键词:
- 抗独特型纳米抗体 /
- SpyCatcher/SpyTag /
- 真菌毒素 /
- 玉米赤霉烯酮 /
- 脱氧雪腐镰刀菌烯醇
Abstract: To improve the immunoassay performance of mycotoxin anti-idiotype nanobody and to realize mimicry of bispecific detected antigens, in this study, anti-idiotype nanobody against zearalenone (ZEN) and deoxynivalenol (DON) were used, and the SpyCatcher/SpyTag tags were fusion-expressed with N28 and Z6, respectively. Based on the in vitro self-assembly, the construction study of bivalent and bispecific anti-idiotype nanobody of N28 and Z6 were carried out. The results showed that the prokaryotic soluble expression of four SpyCatcher/SpyTag labeled fusion anti-idiotype nanobody, N28-SpyCatcher, N28-SpyTag, Z6-SpyCatcher, and Z6-SpyTag were achieved. The IC50 of the competition inhibition standard curves established based on Z6-SpyCatcher and Z6-SpyTag were 0.18 and 0.20 ng/mL, respectively. Their sensitivities were significantly higher than that of the IC50 of monomeric Z6 (0.28 ng/mL). The fusion of the SpyCatcher tag with N28 was favorable for the enhancement of the sensitivity of the N28 assay, with the IC50 value decreased from 94.39 to 42.33 ng/mL, whereas the fusion of SpyTag label with N28 inhibited its binding to DON monoclonal antibody. In addition, bivalent and bispecific anti-idiotype nanobody Z6-Z6, N28-28, and N28-Z6, Z6-N28 of Z6 and N28 were successfully prepared based on the in vitro self-assembly property of SpyCatcher/SpyTag under the simple reaction condition of 4 ℃ for 12 h. The characterization results of the ELISA showed that the bivalent anti-idiotype nanobody retained their binding properties to the corresponding antibodies. The binding properties of the corresponding antibodies, in which N28-Z6 also achieved the binding activity with ZEN and DON antibodies simultaneously, and Z6-N28 only retained the binding activity with ZEN antibody because the binding of SpyTag tag to N28 would hinder its binding to DON antibody. In conclusion, the present study realized a simple and rapid preparation of ZEN/DON bivalent and bispecific anti-idiotype nanobody based on SpyCatcher/SpyTag and suggested that the SpyCatcher tag had a facilitating effect on the immunoassay performance enhancement of ZEN/DON anti-idiotype nanobody.-
Keywords:
- anti-idiotype nanobody /
- SpyCatcher/SpyTag /
- mycotoxin /
- zearalenone /
- deoxynivalenol
-
真菌毒素(Mycotoxin)是由真菌产生的一大类次生代谢产物,常污染于谷物粮食、坚果、水果及其衍生食品中[1],已被证实具有致畸、致癌毒性,目前已成为影响我国食品质量安全的主要因素之一[2−3]。免疫分析方法由于具有较高的灵敏度和特异性,易操作、低成本等特点,被广泛应用真菌毒素的快速检测领域[4−6]。由于真菌毒素的半抗原结构特点,目前其免疫分析方法主要以竞争型为主[5,7],除了需要制备特异好、灵敏度高的单克隆抗体外,还需以真菌毒素半抗原标准品为原料制备检测抗原(全抗原)以实现检测。根据竞争型免疫分析体系的基本原理,其分析性能不仅依赖于抗体的质量,检测抗原的构效组成及制备质量对于免疫分析方法的灵敏度及稳定性也具有决定性的影响[8−10]。然而,目前真菌毒素检测抗原的制备主要基于化学合成方法,其存在合成步骤复杂、批次误差大且对环境造成污染等局限性[11−12],此外,真菌毒素天然半抗原的复杂化学结构也使得通过检测抗原改造来提升免疫分析性能面临较大困难。
有鉴于此,研究者基于抗原模拟表位理论,以多肽或抗独特型纳米抗体为核心元件,开展了真菌毒素抗原模拟物的构建研究,将其替代以天然真菌毒素半抗原为原料所制备的检测抗原,并应用于真菌毒素的免疫分析领域,此达到了优良的检测性能[12−14]。此外,由于多肽及抗独特型纳米抗体具备可生物合成的特点,真菌毒素检测抗原的制备路径不仅实现了由化学合成向生物合成的转变,而且为通过定向改造多肽、抗独特型纳米抗体来改变检测抗原的构效关系,从而调节免疫分析性能提供了便利 [15]。例如,贺贞云[16]在获得赭曲霉毒素A(Ochratoxin A,OTA)的多肽抗原模拟表位的基础上,通过构建噬菌体展示二级突变多肽库,获得了多种灵敏度提升的多肽抗原模拟表位;谌远[17]通过对DON的多肽抗原模拟表位进行丙氨酸突变,发现DON抗原模拟表位D6的第5个氨基酸(组氨酸)突变后与抗体的结合能力会发生不同程度改变,这一发现为通过改造检测抗原来提升免疫分析性能提供了实证支持。然而,相对于多肽分子量小、结构简单的特点,抗独特型纳米抗体具有较为复杂的三级结构及组成[18],此外,纳米抗体在经过分子改造后,在生物合成过程中可能会面临形成包涵体、表达量降低或活性减弱等问题[19],因此,目前通过改造抗独特型纳米抗体来提升真菌毒素检测抗原模拟物性能的报道较少。
SpyCatcher/SpyTag是目前常用的蛋白偶联系统,其操作简单、快速、可实现蛋白质的体外自组装[20],目前已有研究报道使用SpyCatcher/SpyTag作为标签对蛋白进行改造后再进行双特异性抗体构建[21−22]。本研究在前期获得以抗独特型纳米抗体为元件的真菌毒素DON、ZEN抗原模拟物的基础上,基于SpyCatcher/SpyTag标签,分别构建带有SpyCatcher、SpyTag标签的DON、ZEN抗独特型纳米抗体,并基于SpyCatcher/SpyTag具备体外自组装的特点,开展DON、ZEN双价、双特异性抗独特型纳米抗体的体外自组装研究,并探究其免疫分析性能的变化情况,为通过改造抗独特型纳米抗体来提升真菌毒素检测抗原模拟物性能的研究提供前期基础。
1. 材料与方法
1.1 材料与仪器
DON抗独特型纳米抗体表达载体pET25b-N28(N28)[23]、ZEN抗独特型纳米抗体表达载体pET25b-Z6(Z6)[24]、含SpyTag、SpyCatcher编码基因载体、ZEN单克隆抗体、DON单克隆抗体、Rosetta感受态细胞 本实验室自制;限制性内切酶Nco Ⅰ、Not Ⅰ 宝日医生物技术(北京)有限公司;T7通用引物F(TAATACGACTCACTATAGGG)、T7通用引物R(TGCTAGTTATTGCTCAGCGG) 生工生物工程(上海)股份有限公司;Ni-NTA琼脂糖凝胶 美国通用电气公司;SEC分子筛层析柱 美国Citiva公司;SYPRO Orange蛋白凝胶染色剂 西格玛奥德里奇(上海)贸易有限公司;三(羟甲基)氨基甲烷、3,3,5,5-四甲基联苯胺(TMB) 上海阿拉丁生化科技股份有限公司;其他试剂与药品均为国产分析纯。
ZWYC-2933 恒温摇床 上海智城分析仪器制造有限公司;B-580酶标分析仪 深圳市松科科技发展有限公司;DYY-6D型电泳仪 北京六一生物科技有限公司;英赛斯Autopre 25D蛋白纯化仪 苏州英赛斯智能科技有限公司;TCA0096 PCR仪 美国Thermo Fisher Scientific公司;CFX Duet荧光定量PCR仪 美国Bio-Rad公司。
1.2 实验方法
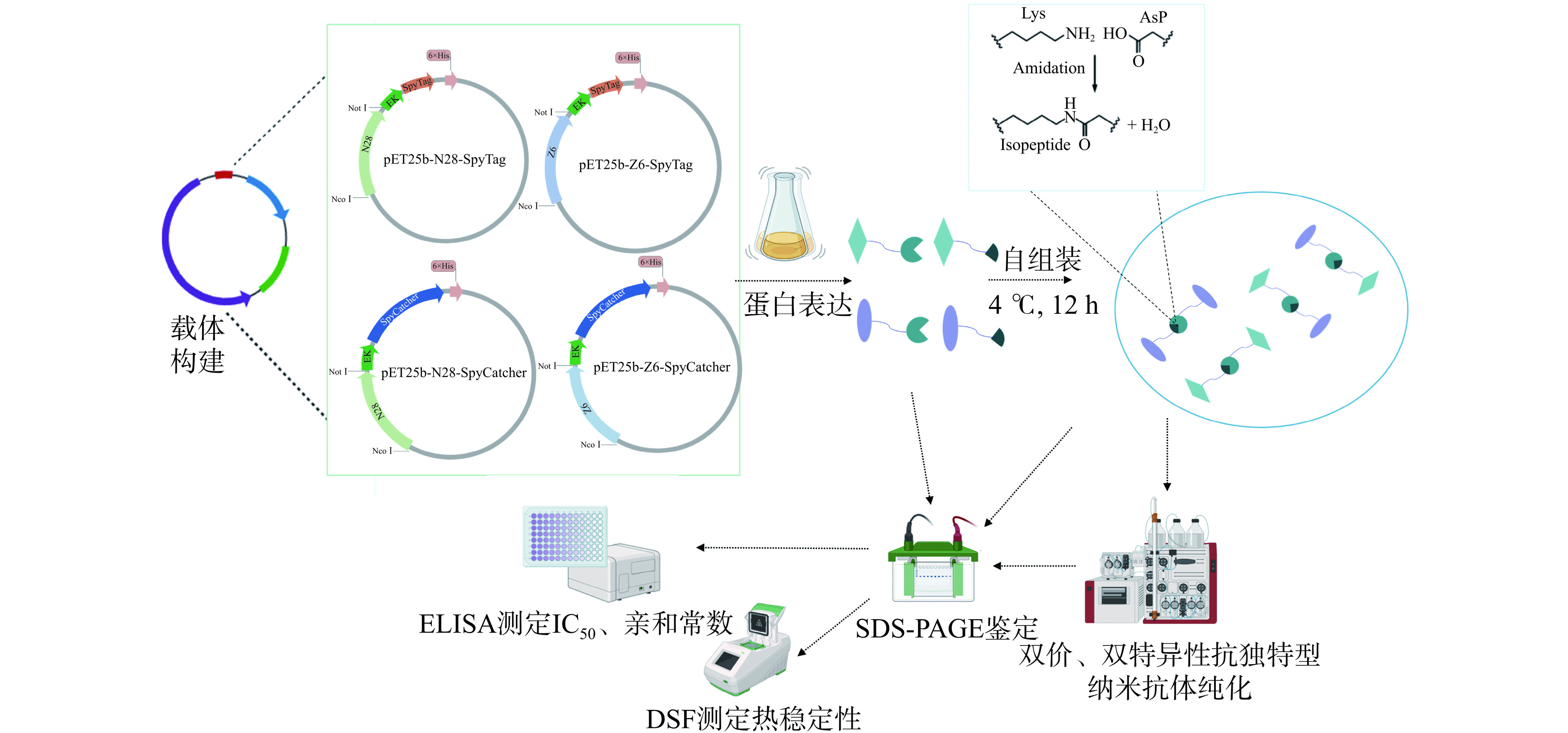
1.2.1 纳米抗体组装方式及实验流程
本研究用到的N28、Z6、SpyTag、SpyCatcher氨基酸序列及具体的双价、双特异性抗独特型纳米抗体组装方式详见表1,本研究的简要流程见图1。
表 1 N28、Z6、SpyTag、SpyCatcher氨基酸序列及双价、双特异性抗独特型纳米抗体图示Table 1. Amino acid sequence of N28, Z6, SpyTag, SpyCatcher and graphical representation of bivalent, bispecific anti-idiotype nanobody名称 氨基酸序列(N-C端) 描述 图示 N28 AQLQLVESGGGLVQAGGSLRLSCAASGRTFSSYGMGWFRQAPGEEREFVAHITRLGVTYYIDSVKGRFAISRDNTENTVYLQMNSLKPEDTAVYYCAARRGSTVPYSANYWSYWGQGTQVTVSSEPKTPKPQ DON抗独特型纳米抗体 
Z6 QVQLVESGGGLVQAGGSLRLSCVASGLTFWTSGMGWFRQAPGKERESVARISWSGGSKFYADSVKGRFTISRDYAKNTVYLQMNSLKPEDTAVYYCAVMEMDIWHEPGYWGQGTQVTVSSEPKTPKPQ ZEN抗独特型纳米抗体 
SpyTag AHIVMVDAYKPTK SpyTag融合标签 
SpyCatcher VDTLSGLSSEQGQSGDMTIEEDSATHIKFSKRDEDGKELAGATMELRDSSGKTISTWISDGQVKDFYLYPGKYTFVETAAPDGYEVATAITFTVNEQGQVTVNGKATKGDAHI SpyCatcher融合标签 
刚性Linker EGKSSGSGESKS 连接肽 
N28-N28 − N28双价抗独特型纳米抗体 
Z6-Z6 − Z6双价抗独特型纳米抗体 
N28-Z6 − N28-Z6双特异性抗独特型
纳米抗体
Z6-N28 − Z6-N28双特异性抗独特型
纳米抗体
注:−表示氨基酸序列未展示。 ![]() 图 1 DON、ZEN双价、双特异性抗独特型纳米抗体的制备及性能鉴定流程图Figure 1. Flowchart for the preparation and performance evaluation of DON, ZEN bivalent, bispecific anti-idiotype nanobody
图 1 DON、ZEN双价、双特异性抗独特型纳米抗体的制备及性能鉴定流程图Figure 1. Flowchart for the preparation and performance evaluation of DON, ZEN bivalent, bispecific anti-idiotype nanobody1.2.2 SpyTag/Catcher标签融合抗独特型纳米抗体的表达
为了实现抗独特型纳米抗体的体外自组装,本研究采用在抗独特型纳米抗体的C端分别融合SpyTag、SpyCatcher标签的方法,基于SpyTag与SpyCatcher可主动识别并自动连接的特性,构建双价或双特异性抗独特型纳米抗体。
1.2.2.1 构建SpyTag/Catcher标签融合抗独特型纳米抗体的表达载体
具体步骤为:通过Nco Ⅰ、Not Ⅰ双酶切获取pET25b-SpyTag、pET25b-SpyCatcher载体上的SpyTag、SpyCatcher基因,反应条件为37 ℃,30 min,酶切产物回收后,16 ℃酶连16 h以将其克隆至N28或Z6的C端,SpyTag/SpyCatcher序列与抗独特型纳米抗体之间以刚性linker进行连接(图1),进而构建4种标签融合抗独特型纳米抗体表达载体,分别为命名为:pET25b-N28-SpyTag(N28-SpyTag)、pET25b-N28-SpyCatcher(N28-SpyCatcher)、pET25b-Z6-SpyTag(Z6-SpyTag)、pET25b-Z6-SpyCatcher(Z6-SpyCatcher)。采用T7通用引物对上述载体的转化子进行菌液PCR验证。PCR扩增条件:预变性95 ℃ 5 min,变性95 ℃ 30 s、退火55 ℃ 20 s、延伸72 ℃ 90 s,共25个循环,终延伸72 ℃ 10 min。将阳性转化子进行DNA测序验证。
1.2.2.2 标签融合抗独特型纳米抗体表达
将构建好的表达载体分别转化至大肠杆菌Rosetta感受态细胞中,0.5 mmol/L异丙基-β-D-硫代半乳糖苷(Isopropyl-beta-D-thiogalactopyranoside,IPTG)、25 ℃条件下诱导表达12 h,离心收集菌体,经超声破碎后取上清过镍柱纯化,用含有20 mmol/L咪唑的缓冲溶液依次清洗柱体,洗去杂蛋白,分别取含有50、100、200 mmol/L咪唑的缓冲液洗脱目的蛋白后用1×PBS 4 ℃过夜透析,各取透析后蛋白10 μL,加入Loading Buffer后采用SDS-PAGE验证纯化产物,通过BCA法进行蛋白质浓度测定(y=1.555x+0.0229,R2=0.9955,y为562 nm处对应的吸光度,x为蛋白浓度)。
1.2.3 抗独特型纳米抗体的体外自组装
1.2.3.1 DON或ZEN双价抗独特型纳米抗体的体外自组装
参考闫志华等[25]的方法并稍作修改,具体步骤为:首先取1.2.1纯化所得N28-SpyTag、N28-SpyCatcher、Z6-SpyTag、Z6-SpyCatcher四种标签融合抗独特型纳米抗体稀释到10 μmol/L,固定反应温度为4 ℃,反应时间为12 h,通过优化摩尔比的方法确定最佳反应摩尔比。为了形成N28-N28或Z6-Z6,按照SpyTag:SpyCatcher不同摩尔比例(1:1、1:2、1:4、1:6、1:8、2:1、4:1、6:1、8:1)取N28-SpyTag与N28-SpyCatcher或Z6-SpyTag与Z6-SpyCatcher于4 ℃下混合反应12 h,终止反应后通过 SDS-PAGE确定体外自组装效率,以选择最佳反应摩尔比。
1.2.3.2 DON、ZEN双特异性抗独特型纳米抗体的体外自组装
N28-Z6、Z6-N28双特异性抗独特型纳米抗体的体外自组装步骤基本相同,不同的是N28-Z6中参与体外自组装反应的抗独特型纳米抗体为N28-SpyCatcher与Z6-SpyTag;Z6-N28中参与体外自组装反应的抗独特型纳米抗体为Z6-SpyCatcher与N28-SpyTag。确定组装摩尔比后将反应浓度增大到50 μmol/L,扩大反应体系至500 μL,以获取更高浓度双特异性抗独特型纳米抗体进行后续鉴定。
1.2.3.3 双价/双特异性抗独特型纳米抗体分离
对于未反应完全的双价/双特异性抗独特型纳米抗体采用体积排阻色谱(Size exclusion chromatography,SEC)分子筛层析柱进行分离。将蛋白溶液用0.22 μm滤膜过滤后用2个柱体积的平衡缓冲液(0.02 mol/L磷酸钠、0.5 mol/L氯化钠,pH=7.4)平衡柱子后进行上样,继续冲洗2个柱体积,不同分子量蛋白在不同时间洗脱出峰,收集所有洗脱峰进行SDS-PAGE 电泳验证,用BCA法进行蛋白质浓度测定,计算公式同1.2.2.2。
1.2.4 ELISA鉴定抗独特型纳米抗体的灵敏度
采用间接竞争ELISA(Enzyme linked immunosorbent assay,ELISA)方法鉴定8种改造后的抗独特型纳米抗体(N28-SpyCatcher、N28-SpyTag、Z6-SpyTag、Z6-SpyCatcher、N28-N28、Z6-Z6、N28-Z6、Z6-N28)的灵敏度。
1.2.4.1 方阵滴定法确定8种抗独特型纳米抗体最佳包被浓度
将8种抗独特型纳米抗体从600 nmol/L开始梯度包被,共包被8个梯度,37 ℃,2 h后用5%脱脂奶粉封闭,封闭结束后添加不同浓度ZEN或DON单克隆抗体(从3 μg/mL开始梯度稀释,稀释8个梯度),后续实验步骤同参考文献[17]。
1.2.4.2 间接竞争ELISA的方法制备竞争抑制标准曲线
根据1.2.3.1中确定的包被浓度将抗独特型纳米抗体进行包被,封闭洗板后每孔加入50 μL梯度稀释的ZEN标准品(50 ng/mL开始稀释)或DON(4000 ng/mL开始稀释,最终梯度为4000、3000、2000、1500、1000、500、250、125、62.5、31.25、15.63、7.81、3.91、1.95、0.98、0.4899、0.25 ng/mL),再加入对应浓度DON或ZEN单克隆抗体;后续测定具体步骤同参考文献[17]。最终结果以结合率(B/B0)=[(竞争抗原孔OD450值−空白对照OD450值)]/非竞争抗原孔为纵坐标,标品浓度为横坐标,使用GraphPad Prism 8拟合绘制竞争抑制曲线。
1.2.5 抗独特型纳米抗体的亲和常数鉴定
采用谌远报道的方法[17],通过间接竞争ELISA法鉴定8种改造后抗独特型纳米抗体(N28-SpyCatcher、N28-SpyTag、Z6-SpyTag、Z6-SpyCatcher、N28-N28、Z6-Z6、N28-Z6、Z6-N28)的亲和常数,具体步骤为:将上述抗独特型纳米抗体包被于96孔酶标板上(200 nmol/L),加入不同浓度ZEN或DON单克隆抗体,抗体从30 μg/mL开始梯度稀释,稀释16个浓度梯度,后续实验步骤同参照文献[17],实验结果以DON单克隆抗体、ZEN单克隆抗体的摩尔浓度作为横坐标,以OD450值为纵坐标绘制反应曲线,并通过GraphPad Prism 8非线性拟合得出IC50,以IC50倒数计算出亲和常数(L/moL)。
1.2.6 抗独特型纳米抗体的热稳定性鉴定
利用差示扫描荧光(Differential scanning fluorimetry,DSF)对标签融合抗独特型纳米抗体、双价/双特异性抗独特型纳米抗体的热稳定性进行鉴定,具体步骤为:用1×PBS将蛋白荧光染料稀释成1:2备用,置于4 ℃保存;将8种改造后抗独特型纳米抗体(N28-SpyCatcher、N28-SpyTag、Z6-SpyTag、Z6-SpyCatcher、N28-N28、Z6-Z6、N28-Z6、Z6-N28)及原始抗独特型纳米抗体N28、Z6稀释为10 μmol/L;取19 μL抗独特型纳米抗体溶液、1 μL蛋白荧光染料置于八连管中(体系中的荧光染料终浓度为1:40),离心后将八连管置于qPCR仪中进行检测,检测条件设置如下:选择溶解曲线、连续模式,扫描温度范围为25~95 ℃,升温速率为0.5 ℃/min,25 ℃平衡4 min,在升温过程中采集数据,报告基团选择HEX,反应体积20 μL。
1.3 数据处理
所有数据均为3次平行实验结果,数据统计分析采用GraphPad Prism 8进行,对数据进行非线性回归分析。
2. 结果与分析
2.1 SpyCatcher/SpyTag标签融合抗独特型纳米抗体的重组表达
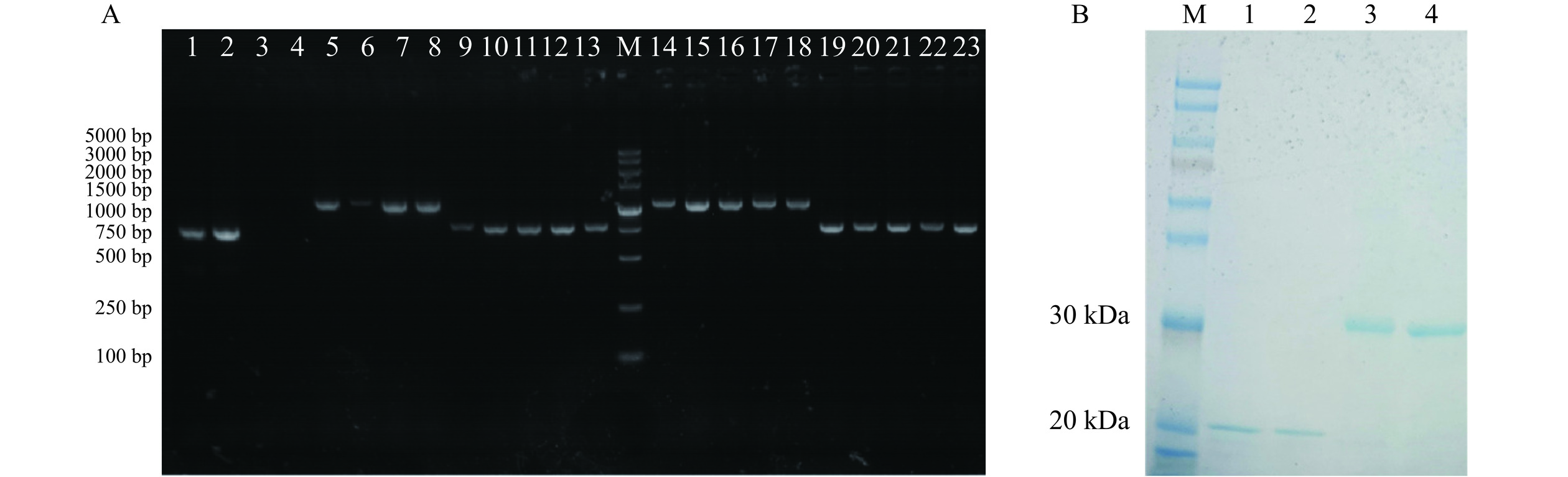
如图2所示,菌液PCR结果表明N28-SpyTag、Z6-SpyTag在750 bp处,N28-SpyCatcher 、Z6-SpyCatcher在1000 bp处出现了目的PCR条带,与预期结果相吻合;测序结果正确后将四种重组表达载体质粒钙转至大肠杆菌宿主菌Rosetta中诱导表达,镍柱纯化后得到目的蛋白,经SDS-PAGE 鉴定分析,结果如图2所示。标签融合抗独特型纳米抗体N28-SpyTag、Z6-SpyTag在22 kDa处,N28-SpyCatcher、Z6-SpyCatcher在33 kDa处出现了单一条带,与预期分子量一致,且均为可溶性表达,纯度大于90%,表明实现了N28、Z6与SpyCatcher/SpyTag标签的可溶性表达。采用BCA蛋白浓度试剂盒进行浓度测定,结果显示N28-SpyTag、N28-SpyCatcher 、Z6-SpyTag、Z6-SpyCatcher的表达量分别约为2.7、15、3.2、13 mg/L。与SpyTag标签相比,抗独特型纳米抗体与SpyCatcher进行融合表达后,其表达量均更高,表明SpyCatcher标签对于纳米抗体具有促溶作用,结果与王琨等[26]的前期研究结论一致。
![]() 图 2 SpyCatcher/SpyTag标签融合抗独特型纳米抗体的PCR鉴定(A)及SDS-PAGE分析(B)注:A图中1为Z6-Nb质粒,2为D28-Nb质粒,3为空白对照(模板为水),4~8为N28-SpyCatcher转化子,M为DL5000,9~13为N28-SpyTag转化子,14~18为Z6-SpyTag转化子,19~23为Z6-SpyCatcher转化子;B图中1为N28-SpyTag,2为Z6-SpyTag,3为N28-SpyCatcher,4为Z6-SpyCatcher,M为蛋白Marker(8~200 kDa)。Figure 2. PCR verification (A) and SDS-PAGE analysis (B) of anti-idiotype nanobody fused with SpyCatcher/SpyTag
图 2 SpyCatcher/SpyTag标签融合抗独特型纳米抗体的PCR鉴定(A)及SDS-PAGE分析(B)注:A图中1为Z6-Nb质粒,2为D28-Nb质粒,3为空白对照(模板为水),4~8为N28-SpyCatcher转化子,M为DL5000,9~13为N28-SpyTag转化子,14~18为Z6-SpyTag转化子,19~23为Z6-SpyCatcher转化子;B图中1为N28-SpyTag,2为Z6-SpyTag,3为N28-SpyCatcher,4为Z6-SpyCatcher,M为蛋白Marker(8~200 kDa)。Figure 2. PCR verification (A) and SDS-PAGE analysis (B) of anti-idiotype nanobody fused with SpyCatcher/SpyTag2.2 抗独特型纳米抗体的体外自组装
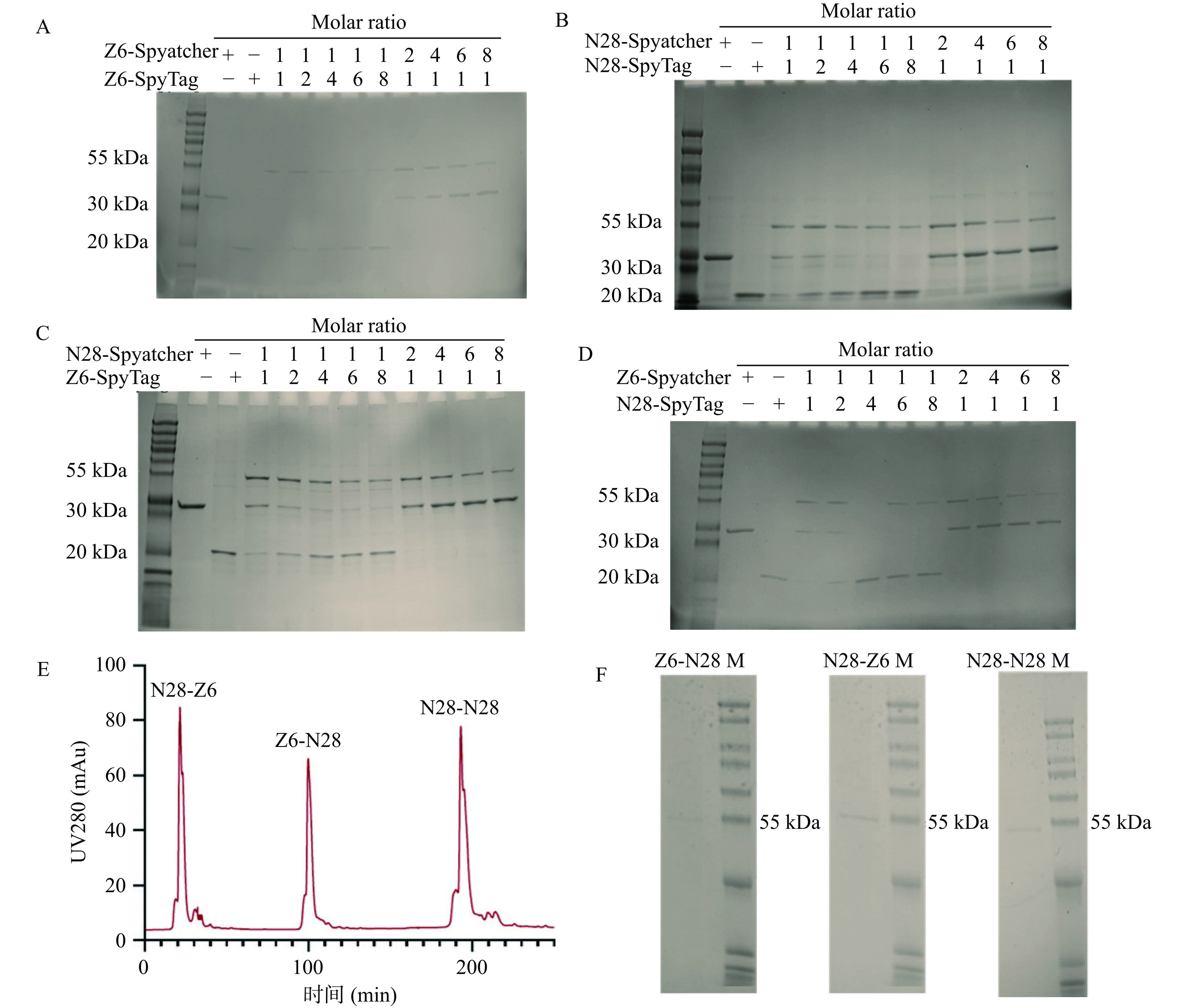
基于SpyCatcher/SpyTag蛋白连接系统的ZEN、DON双价/双特异性抗独特型纳米抗体自组装具有操作简单,反应温和的特点[27]。本研究首先以制备成功的SpyCatcher/SpyTag标签融合抗独特型纳米抗体为元件进行双价抗独特型纳米抗体体外自组装。研究结果(图3A)表明,对于Z6-Z6双价抗独特型纳米抗体的体外自组装,当4 ℃反应12 h、SpyTag:SpyCatcher的反应摩尔比为1:1时,Z6-SpyCatcher与Z6-SpyTag反应趋于完全,并自组装产生约55 kDa大小的Z6-Z6双价抗独特型纳米抗体,而当摩尔比为1:2至1:8时显示Z6-SpyTag过量;摩尔比为2:1~8:1时显示Z6-SpyCatcher过量。
![]() 图 3 SDS-PAGE及SEC纯化鉴定抗独特型纳米抗体的体外自组装产物注:A. Z6-SpyCatcher、Z6-SpyTag两种蛋白自组装;B. N28-SpyCatcher、N28-SpyTag两种蛋白自组装;C. N28-SpyCatcher、Z6-SpyTag两种蛋白自组装;D. Z6-SpyCatcher、N28-SpyTag两种蛋白自组装;E、F分别为Z6-N28、N28-Z6、N28-N28凝胶过滤层析及SDS-PAGE结果;+表示添加该蛋白,−表示未添加该蛋白;Marker(8~200 kDa)。Figure 3. Identification of in vitro self-assembly products of anti-idiotype nanobody by SDS-PAGE and SEC purification
图 3 SDS-PAGE及SEC纯化鉴定抗独特型纳米抗体的体外自组装产物注:A. Z6-SpyCatcher、Z6-SpyTag两种蛋白自组装;B. N28-SpyCatcher、N28-SpyTag两种蛋白自组装;C. N28-SpyCatcher、Z6-SpyTag两种蛋白自组装;D. Z6-SpyCatcher、N28-SpyTag两种蛋白自组装;E、F分别为Z6-N28、N28-Z6、N28-N28凝胶过滤层析及SDS-PAGE结果;+表示添加该蛋白,−表示未添加该蛋白;Marker(8~200 kDa)。Figure 3. Identification of in vitro self-assembly products of anti-idiotype nanobody by SDS-PAGE and SEC purification对于N28-N28双价抗独特型纳米抗体而言,4 ℃反应12 h后产生了N28-N28的产物条带,但是均存在反应物未完全消耗的现象(图3B),后对反应产物采用SEC进行分离,SDS-PAGE结果表明分离后获得了纯度较高的N28-N28双价抗独特型纳米抗体,可用于后续检测(图3F)。与N28-N28双价抗独特型纳米抗体自组装的结果类似,4 ℃反应12 h后均产生了对应的双特异性抗独特型纳米抗体Z6-N28及N28-Z6,但是均存在反应物未完全消耗的现象(图3C、图3D),同样采用SEC进行分离、纯化获得纯度较高的N28-Z6、Z6-N28的双特异性抗独特型纳米抗体(图3E、图3F)。
2.3 ELISA鉴定抗独特型纳米抗体的特异性及灵敏度
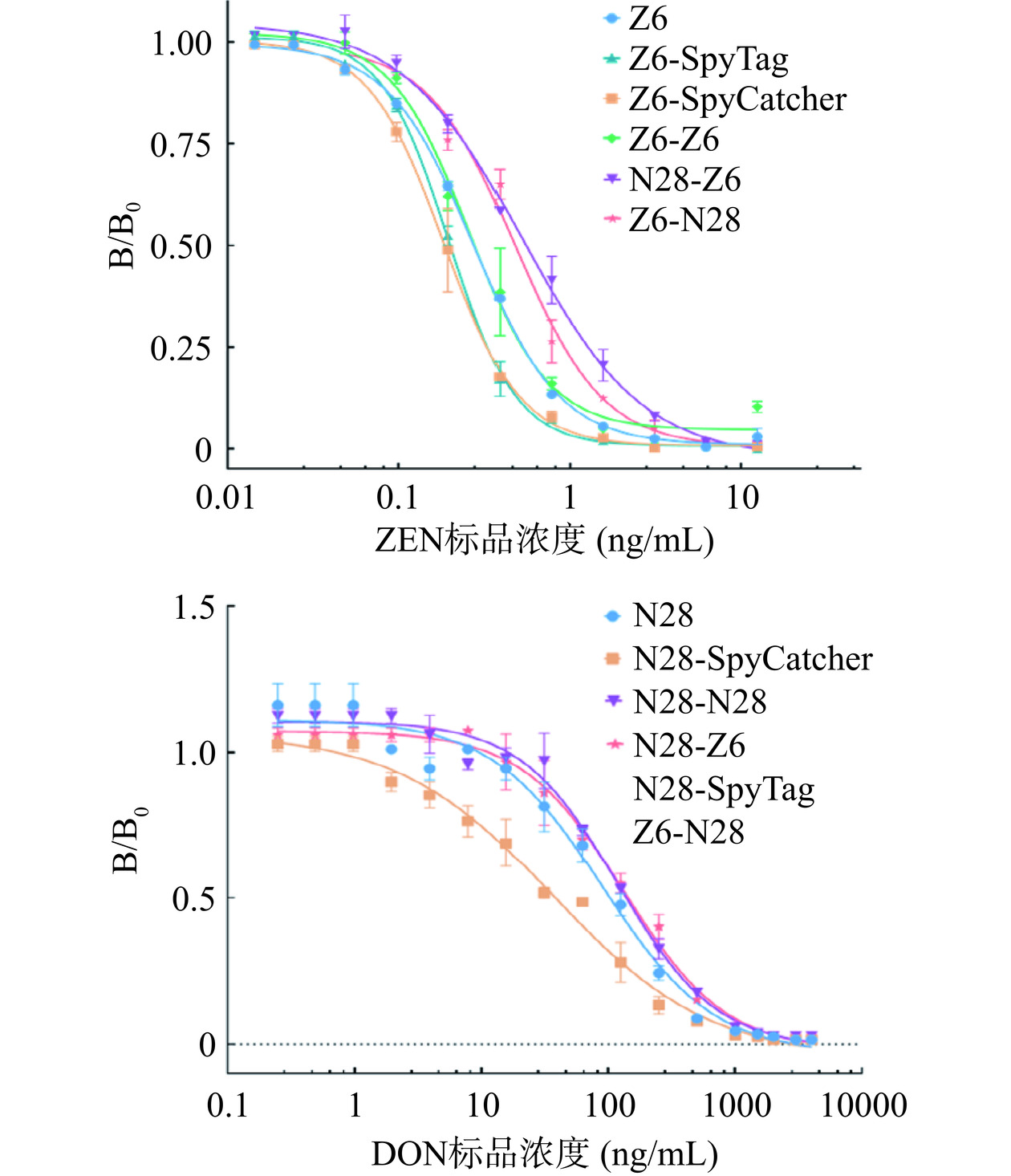
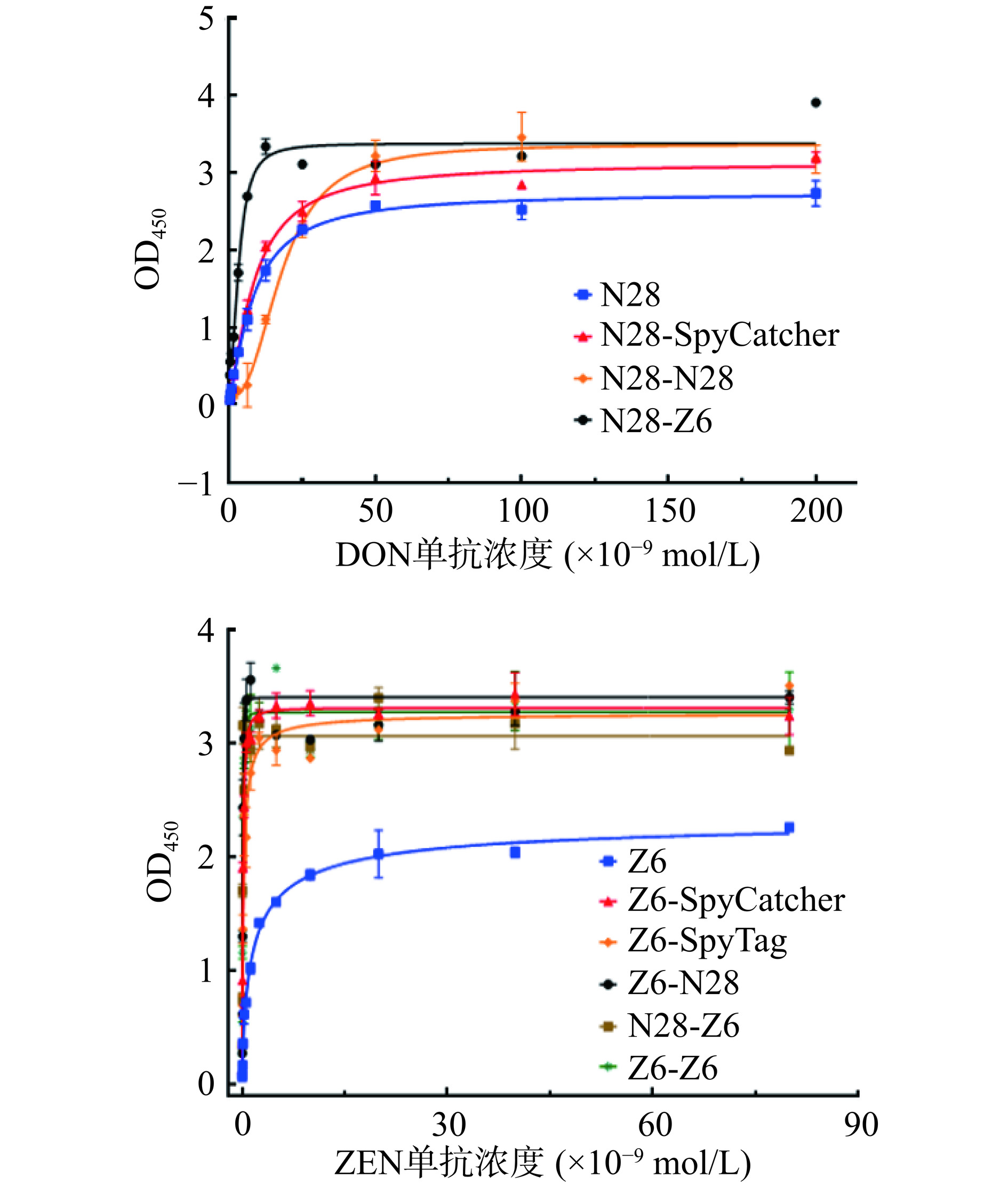
ELISA结果(图4、表2)显示:间接竞争ELISA标准曲线的结果表明,当反应体系中ZEN的浓度增加,反应孔中的OD450呈梯度下降的趋势,显示出竞争结合效应,基于Z6-SpyTag、Z6-SpyCatcher、Z6-Z6所建立的竞争抑制标准曲线的IC50分别为0.20、0.18及0.27 ng/mL,显示改造后的Z6抗独特型纳米抗体的灵敏度均高于改造前的Z6(IC50=0.28 ng/mL)。
![]() 图 4 基于抗独特型纳米抗体的间接竞争ELISA标准曲线Figure 4. Indirect competition ELISA standard curve based on anti-idiotype nanobody表 2 抗独特型纳米抗体IC50汇总Table 2. Summary of IC50 for anti-idiotype nanobody
图 4 基于抗独特型纳米抗体的间接竞争ELISA标准曲线Figure 4. Indirect competition ELISA standard curve based on anti-idiotype nanobody表 2 抗独特型纳米抗体IC50汇总Table 2. Summary of IC50 for anti-idiotype nanobody抗独特型纳米抗体 IC50(ng/mL) Z6 0.28 N28 94.39 Z6-SpyTag 0.20 Z6-SpyCatcher 0.18 N28-SpyTag − N28-SpyCatcher 42.33 Z6-Z6 0.27 N28-N28 122.1 Z6-N28 − Z6-N28 0.48 N28-Z6 0.27 N28-Z6 133.5 注:“−”表示IC50未测出。 然而对于抗独特型纳米抗体N28,ELISA结果显示改造后的抗独特型纳米抗体N28-SpyTag与DON单克隆抗体结合活性变弱,且结合无法被竞争抗原DON阻断,提示SpyTag的氨基酸序列可能对于N28与DON抗体的识别与结合具有干扰作用;而N28-SpyCatcher、N28-N28仍保留了与DON抗体的特异性结合功能,且基于N28-SpyCatcher的间接竞争ELISA灵敏度(IC50=42.33 ng/mL)还高于改造前的N28(IC50=94.39 ng/mL),由于N28-28双价抗独特型纳米抗体是依赖于SpyTag与SpyCatcher的连接,可能是受SpyTag序列的影响,N28-28的灵敏度有所降低(IC50=122.10 ng/mL)。
双特异性抗独特型纳米抗体N28-Z6由N28-SpyCatcher与Z6-SpyTag自组装而成,其保留了对ZEN、DON单克隆抗体结合的特异性,但N28-Z6抗独特型纳米抗体的灵敏度(IC50=133.50 ng/mL)、(IC50=0.27 ng/mL)均低于改造前的N28(IC50,94.39 ng/mL)、Z6(IC50=0.28 ng/mL)。Z6-N28是由Z6-SpyCatcher与N28-SpyTag自组装而成,保留了对ZEN单克隆抗体结合的特异性,但检测灵敏度(IC50,0.48 ng/mL)低于改造前Z6;而由于受N28-SpyTag这一自组装元件的影响,Z6-N28与DON单克隆抗体结合活性变弱,且结合也无法被竞争抗原DON阻断。双价及双特异性抗独特型纳米抗体灵敏度检测结果表明组装元件的原始检测效果对于后续双特异性检测灵敏度有着决定性的影响作用。
2.4 抗独特型纳米抗体的亲和常数鉴定
本研究通过间接ELISA测定SpyCatcher/SpyTag标签融合抗独特型纳米抗体、通过体外自组装获取的双价、双特异性抗独特型纳米抗体的亲和常数,绘制亲和常数拟合曲线。拟合结果显示(图5,表3),对于抗独特型纳米抗体Z6(6.68×108 L/mol)而言,Z6-SpyCatcher(7.61×109 L/mol)、Z6-SpyTag(6.75×109 L/mol)、Z6-Z6(9.11×109 L/mol)的亲和常数均得到了至少一个数量级的提升,当Z6-N28、N28-Z6的ELISA检测中投入的单抗为ZEN单克隆抗体时,检测不会受到DON-SpyTag的影响,因此亲和常数表现为明显提高。表明对于抗独特型纳米抗体Z6来说,对其进行单独的标签融合表达及多聚化改造均能提升其与单克隆抗体结合能力。
![]() 图 5 间接ELISA测定抗独特型纳米抗体的亲和常数Figure 5. Indirect ELISA to determine the affinity of anti-idiotype nanobody表 3 独特型纳米抗体的亲和常数汇总Table 3. Summary of the affinity for anti-idiotype nanobody
图 5 间接ELISA测定抗独特型纳米抗体的亲和常数Figure 5. Indirect ELISA to determine the affinity of anti-idiotype nanobody表 3 独特型纳米抗体的亲和常数汇总Table 3. Summary of the affinity for anti-idiotype nanobody抗独特型纳米抗体 Ka(L/mol) N28 1.18×108 Z6 6.68×108 Z6-SpyTag 7.61×109 Z6-SpyCatcher 6.75×109 N28-SpyTag − N28-SpyCatcher 1.15×108 Z6-Z6 9.11×109 N28-N28 5.52×107 Z6-N28 − Z6-N28 9.43×109 N28-Z6 1.26×1010 N28-Z6 2.79×108 注:“−”表示亲和常数未测出。 对于抗独特型纳米抗体N28(1.18×108 L/mol)而言,N28-SpyCatcher(1.15×108 L/mol)、N28-Z6(2.79×108 L/mol)的亲和常数变化不大:N28-SpyTag与DON单克隆抗体结合活性变弱,且N28-SpyTag参与自组装反应生成的N28-N28的亲和常数下降了一个数量级(5.52×107 L/mol),Z6-N28与DON 单克隆抗体结合活性变弱,无法拟合出亲和常数;再次证明了SpyTag的氨基酸序列对于N28与DON抗体的识别与结合具有干扰作用。
2.5 抗独特型纳米抗体的热稳定性鉴定
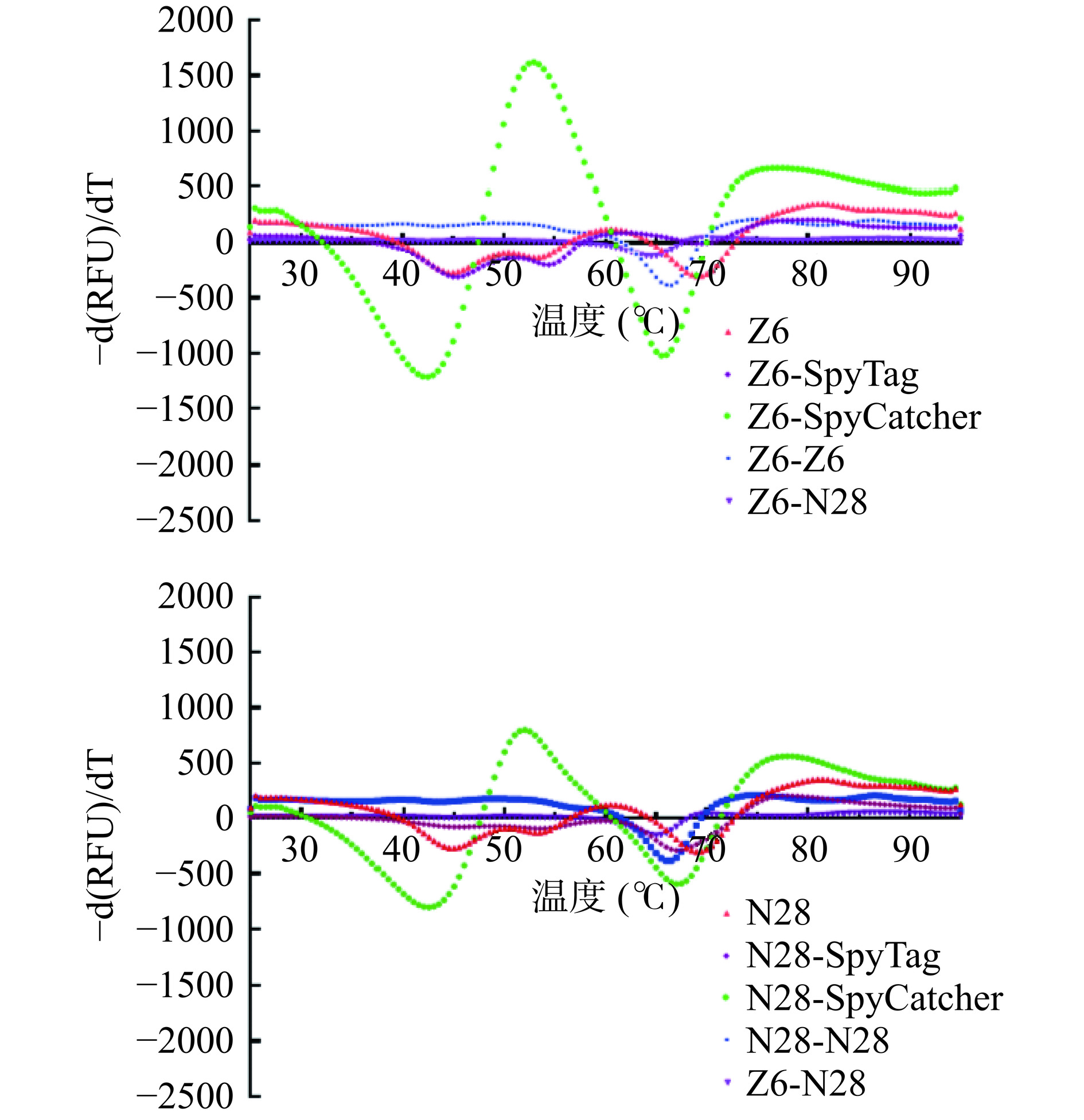
对于Tm的考察可以反映蛋白质的结构稳定性,也能一定程度上评估蛋白空间结构的正确性。当蛋白质热稳定性越强,发生热变性需要的温度就越高,Tm能反映变温过程中蛋白质构象改变的趋势,是衡量蛋白质热稳定性的一个重要指标。本研究采用DSF对单独的N28、Z6抗独特型纳米抗体及SpyCatcher/SpyTag标签融合抗独特型纳米抗体、通过体外自组装获取的双价、双特异性抗独特型纳米抗体进行Tm值测定,结果见图6,表4。对于抗独特型纳米抗体Z6、N28来说,融合了SpyCatcher和SpyTag标签抗独特型纳米抗体的Tm1和Tm2均出现了不同程度的降低,其中添加了SpyCatcher标签的抗独特型纳米抗体Tm值明显降低,Z6-SpyCatcher、N28-SpyCatcher Tm1与Z6、N28相比下降了3 ℃左右;而添加了SpyTag标签的抗独特型纳米抗体Tm值基本没有变化;说明对于抗独特型纳米抗体而言,添加标签会降低其热稳定性,且标签越大,影响越明显。
![]() 图 6 DSF测定抗独特型纳米抗体的荧光曲线一阶导数图Figure 6. First-order derivatives of fluorescence curves for DSF characterization of anti-idiotype nanobody表 4 抗独特型纳米抗体Tm值汇总Table 4. Summary of Tm for anti-idiotype nanobody
图 6 DSF测定抗独特型纳米抗体的荧光曲线一阶导数图Figure 6. First-order derivatives of fluorescence curves for DSF characterization of anti-idiotype nanobody表 4 抗独特型纳米抗体Tm值汇总Table 4. Summary of Tm for anti-idiotype nanobody抗独特型纳米抗体 Tm1(℃) Tm2(℃) N28 42.6 65 Z6 45 70 Z6-SpyTag 42.4 69 Z6-SpyCatcher 43 65 N28-SpyTag 45 69 N28-SpyCatcher 45 67 Z6-Z6 35 N28-N28 46 60 Z6-N28 45 59 N28-Z6 35.8 Z6-N28 35.3 对体外自组装所得双价、双特异性抗独特型纳米抗体进行Tm值测定,实验结果表明双价及双特异性抗独特型纳米抗体组装完成后只有一个Tm值,且与原始Z6或N28相比,Z6-Z6、N28-N28、Z6-N28、N28-Z6的Tm值均出现了降低,其中,Z6-N28 Tm值变化最明显,下降了5 ℃。猜测SpyCatcher和SpyTag反应生成异肽键后受温度影响较小,在较低温度时不会发生去折叠,因此只有一个Tm值,但其存在也会影响抗独特型纳米抗体的热稳定性。
3. 讨论与结论
有研究报道指出,抗独特型抗体作为包被抗原时,检测灵敏度得到了明显提升[28−30]。其中,抗独特型纳米抗体由于特殊的分子结构更适于用作真菌毒素免疫检测抗原模拟物[11]。然而目前如何对抗独特型纳米抗体进行改造以提高提免疫分析检测性能的相关报道较少,其免疫分析性能变化规律及机制仍未知。此外,从真菌毒素免疫检测抗原模拟物的角度出发,实现真菌毒素的多重检测也具有重要意义。
本研究首先将SpyCatcher/SpyTag与Z6、N28进行融合表达,并对融合后蛋白进行多种手段表征,实验结果表明SpyCatcher是一种较好的促溶蛋白,与其进行连接后Z6、N28的活性及检测灵敏度均得以提升。而N28与SpyTag融合后活性下降明显,Z6与SpyTag融合后结果却恰恰相反,其活性明显增强,该现象可能与两种纳米抗体活性位点相关,后续可通过改变连接的Linker种类、将SpyTag融合至纳米抗体N端手段探寻其活性变化规律。在进行标准曲线绘制时发现添加标准品的孔OD值比未加标准品的0值孔要稍大,出现跳孔现象,且主要是在添加了低浓度的标准品的情况下,推测可能是在低浓度的标准品下,其竞争抑制效果不佳,以及ELISA的系统、操作误差所致。在ELISA体系中,呈现线性阻断的标准品浓度主要集中在一定范围内,过高及过低浓度的标准品所形成的阻断率可能不适合用于定量分析,这种非线性浓度下,B/B0>1的情况在已报道的文献中也有所体现[31−33]。单独的抗独特型纳米抗体在与SpyCatcher/SpyTag标签融合后Tm值降低,说明融合标签具有降低蛋白热稳定性效果,且标签分子量越大,效果越显著。
通过SpyCatcher/SpyTag连接系统构建的ZEN-DON双价、双特异性抗独特型纳米抗体连接后仍保持原有结合特异性,但对于间接竞争ELISA检测灵敏度来说,均出现了一定程度的降低,可能是由于蛋白空间位阻问题,原始抗独特型纳米抗体与其余蛋白融合后将一部分原始蛋白结合位点将被结合。后续可通过分子对接等研究手段对其进行模拟以探究内在规律。组装完的双价及双特异性抗独特型纳米抗体只有一个Tm,较原始Tm均出现了降低,与标签融合蛋白规律一致,进一步揭示了纳米抗体结构的延伸会导致其热稳定性的下降。因此,在进行抗独特型纳米抗体改造时应根据实验需求进行选择。
综上,本研究首先通过将N28、Z6抗独特型纳米抗体与SpyCatcher/SpyTag标签进行融合表达,制备体外自组装元件;随后采用SpyCatcher/SpyTag连接系统实现了DON-ZEN双价、双特异性抗独特型纳米抗体的体外自组装,结果表明在自组装后两种抗独特型纳米抗体特异性并未发生改变,均保持了各自的原有活性,改造过程无需使用化学试剂、反应条件温和、便捷。此外,本研究也证明了SpyCatcher/SpyTag连接系统在相同/不同抗独特型纳米抗体连接方面的通用性,为其它抗独特型纳米抗体的改造提供了借鉴意义。
-
![]()
图 1 DON、ZEN双价、双特异性抗独特型纳米抗体的制备及性能鉴定流程图
Figure 1. Flowchart for the preparation and performance evaluation of DON, ZEN bivalent, bispecific anti-idiotype nanobody
![]()
图 2 SpyCatcher/SpyTag标签融合抗独特型纳米抗体的PCR鉴定(A)及SDS-PAGE分析(B)
注:A图中1为Z6-Nb质粒,2为D28-Nb质粒,3为空白对照(模板为水),4~8为N28-SpyCatcher转化子,M为DL5000,9~13为N28-SpyTag转化子,14~18为Z6-SpyTag转化子,19~23为Z6-SpyCatcher转化子;B图中1为N28-SpyTag,2为Z6-SpyTag,3为N28-SpyCatcher,4为Z6-SpyCatcher,M为蛋白Marker(8~200 kDa)。
Figure 2. PCR verification (A) and SDS-PAGE analysis (B) of anti-idiotype nanobody fused with SpyCatcher/SpyTag
![]()
图 3 SDS-PAGE及SEC纯化鉴定抗独特型纳米抗体的体外自组装产物
注:A. Z6-SpyCatcher、Z6-SpyTag两种蛋白自组装;B. N28-SpyCatcher、N28-SpyTag两种蛋白自组装;C. N28-SpyCatcher、Z6-SpyTag两种蛋白自组装;D. Z6-SpyCatcher、N28-SpyTag两种蛋白自组装;E、F分别为Z6-N28、N28-Z6、N28-N28凝胶过滤层析及SDS-PAGE结果;+表示添加该蛋白,−表示未添加该蛋白;Marker(8~200 kDa)。
Figure 3. Identification of in vitro self-assembly products of anti-idiotype nanobody by SDS-PAGE and SEC purification
![]()
图 4 基于抗独特型纳米抗体的间接竞争ELISA标准曲线
Figure 4. Indirect competition ELISA standard curve based on anti-idiotype nanobody
![]()
图 5 间接ELISA测定抗独特型纳米抗体的亲和常数
Figure 5. Indirect ELISA to determine the affinity of anti-idiotype nanobody
![]()
图 6 DSF测定抗独特型纳米抗体的荧光曲线一阶导数图
Figure 6. First-order derivatives of fluorescence curves for DSF characterization of anti-idiotype nanobody
表 1 N28、Z6、SpyTag、SpyCatcher氨基酸序列及双价、双特异性抗独特型纳米抗体图示
Table 1 Amino acid sequence of N28, Z6, SpyTag, SpyCatcher and graphical representation of bivalent, bispecific anti-idiotype nanobody
名称 氨基酸序列(N-C端) 描述 图示 N28 AQLQLVESGGGLVQAGGSLRLSCAASGRTFSSYGMGWFRQAPGEEREFVAHITRLGVTYYIDSVKGRFAISRDNTENTVYLQMNSLKPEDTAVYYCAARRGSTVPYSANYWSYWGQGTQVTVSSEPKTPKPQ DON抗独特型纳米抗体 Z6 QVQLVESGGGLVQAGGSLRLSCVASGLTFWTSGMGWFRQAPGKERESVARISWSGGSKFYADSVKGRFTISRDYAKNTVYLQMNSLKPEDTAVYYCAVMEMDIWHEPGYWGQGTQVTVSSEPKTPKPQ ZEN抗独特型纳米抗体 SpyTag AHIVMVDAYKPTK SpyTag融合标签 SpyCatcher VDTLSGLSSEQGQSGDMTIEEDSATHIKFSKRDEDGKELAGATMELRDSSGKTISTWISDGQVKDFYLYPGKYTFVETAAPDGYEVATAITFTVNEQGQVTVNGKATKGDAHI SpyCatcher融合标签 刚性Linker EGKSSGSGESKS 连接肽 N28-N28 − N28双价抗独特型纳米抗体 Z6-Z6 − Z6双价抗独特型纳米抗体 N28-Z6 − N28-Z6双特异性抗独特型
纳米抗体Z6-N28 − Z6-N28双特异性抗独特型
纳米抗体注:−表示氨基酸序列未展示。  下载: 导出CSV
下载: 导出CSV
表 2 抗独特型纳米抗体IC50汇总
Table 2 Summary of IC50 for anti-idiotype nanobody
抗独特型纳米抗体 IC50(ng/mL) Z6 0.28 N28 94.39 Z6-SpyTag 0.20 Z6-SpyCatcher 0.18 N28-SpyTag − N28-SpyCatcher 42.33 Z6-Z6 0.27 N28-N28 122.1 Z6-N28 − Z6-N28 0.48 N28-Z6 0.27 N28-Z6 133.5 注:“−”表示IC50未测出。
下载: 导出CSV
表 3 独特型纳米抗体的亲和常数汇总
Table 3 Summary of the affinity for anti-idiotype nanobody
抗独特型纳米抗体 Ka(L/mol) N28 1.18×108 Z6 6.68×108 Z6-SpyTag 7.61×109 Z6-SpyCatcher 6.75×109 N28-SpyTag − N28-SpyCatcher 1.15×108 Z6-Z6 9.11×109 N28-N28 5.52×107 Z6-N28 − Z6-N28 9.43×109 N28-Z6 1.26×1010 N28-Z6 2.79×108 注:“−”表示亲和常数未测出。
下载: 导出CSV
表 4 抗独特型纳米抗体Tm值汇总
Table 4 Summary of Tm for anti-idiotype nanobody
抗独特型纳米抗体 Tm1(℃) Tm2(℃) N28 42.6 65 Z6 45 70 Z6-SpyTag 42.4 69 Z6-SpyCatcher 43 65 N28-SpyTag 45 69 N28-SpyCatcher 45 67 Z6-Z6 35 N28-N28 46 60 Z6-N28 45 59 N28-Z6 35.8 Z6-N28 35.3
下载: 导出CSV
-
[1] CIMBALO A, ALONSO-GARRIDO M, FONT G, et al. Toxicity of mycotoxins in vivo on vertebrate organisms:A review[J]. Food and Chemical Toxicology,2020,137:111161. doi: 10.1016/j.fct.2020.111161
[2] MARZOCCO S, RUSSO R, BIANCO G, et al. Pro-apoptotic effects of nivalenol and deoxynivalenol trichothecenes in J774A. 1 murine macrophages[J]. Toxicology Letters,2009,189(1):21−26. doi: 10.1016/j.toxlet.2009.04.024
[3] 谭莉, 孟繁磊, 范宏, 等. 超高效液相色谱-串联质谱法同时测定玉米中9种真菌毒素[J]. 食品工业科技,2021,42(7):240−245. [TAN L, MENG F L, FAN H, et al. Determination of 9 kinds of mycotoxins in maize by Ultra Performance Liquid Chromatography-tandem Mass Spectrometry[J]. Science and Technology of Food Industry,2021,42(7):240−245.] TAN L, MENG F L, FAN H, et al. Determination of 9 kinds of mycotoxins in maize by Ultra Performance Liquid Chromatography-tandem Mass Spectrometry[J]. Science and Technology of Food Industry, 2021, 42(7): 240−245.
[4] TURNER N W, BRAMHMBHATT H, SZABO-VEZSE M, et al. Analytical methods for determination of mycotoxins:An update (2009~2014)[J]. Analytica Chimica Acta,2015,901:12−33. doi: 10.1016/j.aca.2015.10.013
[5] MILLS C, CAMPBELL K. A new chapter for anti-idiotypes in low molecular weight compound immunoassays[J]. Trends in Biotechnology,2022,40(9):1102−1120. doi: 10.1016/j.tibtech.2022.02.008
[6] 邱雨楼. 纳米抗体N-28模拟DON抗原的分子机制研究及定点改造[D]. 南昌:南昌大学, 2016. [QIU Y L. Understanding molecular mechanism of DON antigen mimicry by nanobody N-28 and its modification by site saturation mutagenesis[D]. Nanchang:Nanchang University, 2016.] QIU Y L. Understanding molecular mechanism of DON antigen mimicry by nanobody N-28 and its modification by site saturation mutagenesis[D]. Nanchang: Nanchang University, 2016.
[7] 舒梅. 抗独特型纳米抗体的亲和力成熟及检测伏马菌素B1绿色免疫分析方法的研究[D]. 南昌:南昌大学, 2016. [SHU M. Affinity maturation of an anti-idiotypic nanobody and studied on environment-friendly immunoassay for determination of fumonisin B1 [D]. Nanchang:Nanchang University, 2016.] SHU M. Affinity maturation of an anti-idiotypic nanobody and studied on environment-friendly immunoassay for determination of fumonisin B1 [D]. Nanchang: Nanchang University, 2016.
[8] SHI L H, YU T, LUO M E, et al. Preparation monoclonal β-type anti-idiotype antibody of zearalenone and development of green ELISA quantitative detecting technique[J]. Preparative Biochemistry & Biotechnology,2020,50(4):419−424.
[9] LIU R R, XU L, QIU X M, et al. An immunoassay for determining aflatoxin B1 using a recombinant phage as a nontoxic coating conjugate[J]. Journal of Food Safety,2012,32(3):318−325. doi: 10.1111/j.1745-4565.2012.00383.x
[10] WANG Y R, WANG H, LI P W, et al. Phage-displayed peptide that mimics aflatoxins and its application in immunoassay[J]. Journal of Agricultural and Food Chemistry,2013,61(10):2426−2433. doi: 10.1021/jf4004048
[11] YU G G, WANG J M, ZHANG Y, et al. Anti-Idiotypic nanobody alkaline phosphatase fusion protein-triggered on–off–on fluorescence immunosensor for aflatoxin in cereals[J]. Journal of Agricultural and Food Chemistry,2023,71(45):17391−17398.
[12] LIU X, XU Y, HE Q H, et al. Application of mimotope peptides of fumonisin B1 in peptide ELISA[J]. Journal of Agricultural and Food Chemistry,2013,61(20):4765−4770. doi: 10.1021/jf400056p
[13] XU Y, CHEN B, HE Q H, et al. New approach for development of sensitive and environmentally friendly immunoassay for mycotoxin fumonisin B1 based on using peptide-MBP fusion protein as substitute for coating antigen[J]. Analytical Chemistry,2014,86(16):8433−8440. doi: 10.1021/ac502037w
[14] SHU M, XU Y, DONG J X, et al. Development of a noncompetitive idiometric nanobodies phage immumoassay for the determination of fumonisin B1[J]. Food and Agricultural Immunology,2019,30(1):510−521. doi: 10.1080/09540105.2019.1604637
[15] XIONG Y, LENG Y K, LI X M, et al. Emerging strategies to enhance the sensitivity of competitive ELISA for detection of chemical contaminants in food samples[J]. TrAC Trends in Analytical Chemistry,2020,126:115861. doi: 10.1016/j.trac.2020.115861
[16] 贺贞云. 赭曲霉毒素A模拟表位二级库的构建及其全抗原的生物合成[D]. 南昌:南昌大学, 2015. [HE Z Y. Second-generation peptide library construction of mimotope of ochratoxin a and biosynthesis of its complete antigen [D]. Nanchang:Nanchang University, 2015.] HE Z Y. Second-generation peptide library construction of mimotope of ochratoxin a and biosynthesis of its complete antigen [D]. Nanchang: Nanchang University, 2015.
[17] 谌远. 呕吐毒素抗原模拟表位的定向突变及其免疫层析试纸条的制备[D]. 南昌:南昌大学, 2022. [SHENG Y. Directed mutation for deoxynivalenol antigenic mimotopes and development of immunochromatographic strips[D]. Nanchang:Nanchang University, 2022.] SHENG Y. Directed mutation for deoxynivalenol antigenic mimotopes and development of immunochromatographic strips[D]. Nanchang: Nanchang University, 2022.
[18] XIONG L, ZHANG X Q, XU Y, et al. Anti-idiotypic VHH mediated environmentally friendly immunoassay for citrinin without mycotoxin[J]. Food and Agricultural Immunology,2020,31(1):968−984. doi: 10.1080/09540105.2020.1795631
[19] VASSEGHIAN Y, MORADI M, DRAGOI E N, et al. A review on mycotoxins detection techniques in edible oils[J]. International Journal of Environmental Analytical Chemistry,2022,102(9):2125−2139. doi: 10.1080/03067319.2020.1750607
[20] GREENWOOD S N, KULKARNI R S, MIKHAIL M, et al. Replication protein a enhances kinetics of uracil DNA glycosylase on ssDNA and across DNA junctions:Explored with a DNA repair complex produced with spycatcher/spytag ligation[J]. ChemBioChem,2023,24(10):e202200765. doi: 10.1002/cbic.202200765
[21] MEI L, ZAPPALA F, TSOURKAS A. Rapid production of bispecific antibodies from off-the-shelf IgGs with high yield and purity[J]. Bioconjugate Chemistry,2021,33(1):134−141.
[22] YUMURA K, AKIBA H, NAGATOISHI S, et al. Use of SpyTag/SpyCatcher to construct bispecific antibodies that target two epitopes of a single antigen[J]. The Journal of Biochemistry,2017,162(3):203−210. doi: 10.1093/jb/mvx023
[23] QIU Y L, HE Q H, XU Y, et al. Deoxynivalenol-mimic nanobody isolated from a naïve phage display nanobody library and its application in immunoassay[J]. Analytica Chimica Acta,2015,887:201−208. doi: 10.1016/j.aca.2015.06.033
[24] WANG X X, HE Q H, XU Y, et al. Anti-idiotypic VHH phage display-mediated immuno-PCR for ultrasensitive determination of mycotoxin zearalenone in cereals[J]. Talanta,2016,147:410−415. doi: 10.1016/j.talanta.2015.09.072
[25] 闫治华, 刘娜, 刘远声, 等. 人工纤维小体骨架蛋白的分子设计与自组装[J]. 中国生物化学与分子生物学报,2022,38(12):1694−1702. [YAN Z H, LIU N, LIU Y S, et al. Molecular design and self-assembly of cellulosomal scaffoldin[J]. Chinese Journal of Biochemistry and Molecular Biology,2022,38(12):1694−1702.] YAN Z H, LIU N, LIU Y S, et al. Molecular design and self-assembly of cellulosomal scaffoldin[J]. Chinese Journal of Biochemistry and Molecular Biology, 2022, 38(12): 1694−1702.
[26] 王锟, 李日飞, 何伟国, 等. SpyCatcher作为一种新的高效促溶标签蛋白的探讨[J]. 解剖科学进展,2017,23(4):356−359. [WANG K, LI R F, HE W G, et al. SpyCatcher as a new high efficient label protein J]. Progress of Anatomical Sciences,2017,23(4):356−359.
[27] DRISCOLL C L, KEEBLE A H, HOWARTH M. SpyCombinator assembly of bispecific binders[J]. bioRxiv, 2023:2023.08. 31.555700.
[28] XU Y, XIONG L, LI Y P, et al. Anti-idiotypic nanobody as citrinin mimotope from a naive alpaca heavy chain single domain antibody library[J]. Analytical and Bioanalytical Chemistry,2015,407:5333−5341. doi: 10.1007/s00216-015-8693-3
[29] ZHANG C X, ZHANG W Q, TANG X Q, et al. Change of amino acid residues in idiotypic nanobodies enhanced the sensitivity of competitive enzyme immunoassay for mycotoxin ochratoxin a in cereals[J]. Toxins,2020,12(4):273. doi: 10.3390/toxins12040273
[30] SHU M, XU Y, WANG D, et al. Anti-idiotypic nanobody:A strategy for development of sensitive and green immunoassay for Fumonisin B1[J]. Talanta,2015,143:388−393. doi: 10.1016/j.talanta.2015.05.010
[31] YANG X, SUN Z C, HE Z Y, et al. Combination of nanobody and peptidomimetic to develop novel immunoassay platforms for detecting ochratoxin A in cereals[J]. Food Chemistry,2023,429:137018. doi: 10.1016/j.foodchem.2023.137018
[32] WANG Y D, QIN J A, ZHANG J, et al. Rapid screening of imidacloprid residue in grains and medicinal herbs:A newly designed hapten and monoclonal antibody[J]. Journal of Pharmaceutical and Biomedical Analysis,2022,219:114931. doi: 10.1016/j.jpba.2022.114931
[33] DAI P, ZHANG Y, HONG Y P, et al. Production of high affinity monoclonal antibody and development of indirect competitive chemiluminescence enzyme immunoassay for gentamicin residue in animal tissues[J]. Food Chemistry,2023,400:134067. doi: 10.1016/j.foodchem.2022.134067
 下载:
下载:






计量
- 文章访问数: 0
- HTML全文浏览量: 0
- PDF下载量: 0
