


Heterogeneity of Spoilage Potential of Meat-borne Bacillus
-
摘要: 为评估芽孢杆菌的致腐潜能异质性,本研究以从腐败低温香肠中分离出来的6 株芽孢杆菌为研究对象,探究其在20 ℃(常温)和37 ℃(最适生长温度)条件下的致腐性能。试验分别测定了6株芽孢杆菌的生长曲线、产酸能力、产气能力、蛋白酶活力、脂肪酶活力和淀粉酶活力。结果表明,蜡样芽孢杆菌(C)的产酸能力较强,在37 ℃条件下培养48 h,pH较空白组下降0.71;解淀粉芽孢杆菌(G)的产蛋白酶和淀粉酶的能力更强,其蛋白酶活力高达7.17 U/mL,37 ℃条件下培养至5 d的淀粉分解圈面积为16.53 cm2;蜡样芽孢杆菌(C)的综合腐败能力最弱,具体表现为其蛋白酶活力为2.63 U/mL,37 ℃条件下培养5 d的淀粉分解圈面积为7.55 cm2。同时发现生长温度对菌株致腐性能有较大影响。研究结果说明芽孢杆菌菌株的致腐性能存在显著的异质性,在探究其腐败特性时,应充分考虑菌株的异质性。Abstract: In order to assessing the heterogeneity of Bacillus sp. isolates spoilage potential, six Bacillus sp. isolates isolated from spoiled low-temperature sausages were used to investigate the heterogeneity of their spoilage properties at 20 ℃ (room temperature) and 37 ℃ (optimum growth temperature). The growth curve, acid production capacity, gas production capacity, protease activity, lipase activity, and amylase activity of six strains of Bacillus sp. were measured in the experiment. The results revealed that Bacillus cereus (C) had the greatest capacity for producing acid and that after 48 hours at 37 ℃, the pH in the culture medium dropped by 0.71 relative to the blank control group. Following a 5-day culture at 37 ℃, Bacillus amyloliquefaciens (G) displayed the greatest protease and amylase activities, with a maximum protease activity of 7.17 U/mL and a decomposition circle area of starch agar of 16.53 cm2. Bacillus cereus (C) exhibited the least comprehensive spoiling ability, as evidenced by its protease activity of 2.63 U/mL and the decomposition circle area of starch agar of 7.55 cm2 after 5 days of culture at 37 ℃. Furthermore, it was discovered that the growth temperature significantly impacted the spoilage potential of the strains. In summary, there was great heterogeneity in the spoilage characteristics of Bacillus sp.. When examining its spoilage traits, it is important to consider this heterogeneity.
-
Keywords:
- meat products /
- Bacillus sp. /
- spoilage potential /
- protease /
- lipase /
- amylase
-
我国肉与肉制品产量巨大,2021年人均肉类消费量达到了23.9 kg[1],产业发展势头迅猛。肉品具有较高的水分活度和适宜的pH,极易被微生物污染引起腐败变质[2]。在肉品加工、储存和零售过程中,微生物是导致肉品腐败的主要原因,每年给肉类行业造成数十亿美元的经济损失[3]。因此,防止或延缓肉品腐败是全球食品行业共同关注的问题,也是近年来食品科学领域的研究热点[4−5]。
芽孢杆菌作为食品中常见的优势腐败菌,广泛地存在于在各类食品中。国内外关于芽孢杆菌的研究较多,对芽孢杆菌的形态、分类等已有较为透彻的研究[6−7],研究证明芽孢杆菌是肉制品中的优势腐败菌之一。李冉等[8]对狮子头中腐败菌进行菌群结构分析,发现芽孢杆菌在三个处理组中所占的丰度值为20%左右,是狮子头腐败过程中的主要优势菌。甯雨荞等[9]在分离市售笋子烧牛肉中的腐败菌时发现枯草芽孢杆菌的致腐能力最强。同时,芽孢杆菌还具有较强的抗逆性[10−12],传统的巴氏杀菌很难将其杀灭,在后续合适的环境条件下会迅速繁殖,造成食品腐败变质[13−14]。WEBB等[15]综述了蜡样芽孢杆菌在低温食品中的危害,杨啸吟等[16]进一步论述了不同包装的冷却肉腐败过程中挥发性气体与芽孢杆菌等腐败菌之间的关联,再次表明芽孢杆菌是导致肉类腐败的主要细菌。目前针对芽孢杆菌的研究已证明其为低温肉制品中腐败能力较强的腐败菌,但缺乏对于肉源性芽孢杆菌不同菌株之间腐败性能异质性的研究。本研究认为,分析不同芽孢杆菌之间腐败性能的差异,是研发针对性防控芽孢杆菌污染技术的基础。
在微生物污染之后,肉品常见的腐败特征包括产酸、产气和发粘等[17],这是由微生物多种内源酶的氧化分解作用引起的。为了探究不同芽孢杆菌在肉品腐败中的主要致腐特性,根据肉品腐败特征,本研究拟探索6株肉源性芽孢杆菌的产酸、产气、腐败酶等特性,为后续进行肉制品中芽孢杆菌针对性防控提供理论基础。
1. 材料与方法
1.1 材料与仪器
试验所用的6 株芽孢杆菌均由南京农业大学国家肉品质量安全控制工程技术研究中心提供,分离自低温香肠,具体见表1;LB营养琼脂、LB肉汤、玉米油(脂酶)细菌微量生化鉴定管、可溶性淀粉琼脂、卢戈氏碘液等 均购于青岛海博生物技术有限公司;干酪素 购于北京索莱宝科技有限公司;磷酸二氢钠、磷酸氢二钠 均购于国药集团化学试剂有限公司;无水乙醇、95%乙醇 均购于上海麦克林生化科技有限公司。
表 1 6株芽孢杆菌信息Table 1. Information of 6 strains of Bacillus编号 拉丁名 中文名 C Bacillus cereus 蜡样芽孢杆菌 D Bacillus safensis 沙福芽孢杆菌 K Bacillus altitudinis 高山芽孢杆菌 E Bacillus safensis 沙福芽孢杆菌 G Bacillus amyloliquefaciens 解淀粉芽孢杆菌 J Bacillus licheniformis 地衣芽孢杆菌 BIO Ⅱ Adcance4 Sterile GARD生物安全柜 西班牙Telstar公司;Scan1200自动影像分析菌落计数仪 法国Interscience公司;DRP-9162型电热恒温培养箱 上海森信实验仪器有限公司;SQL1010C 立式压力蒸汽灭菌器 日本Yamato公司;涡旋振荡器 美国 Scilogex公司;Testo 205便携式pH 计 德国Testo公司;M2e酶标仪 美国MD公司;PD500匀浆机 英国PRIMASCI公司;MUL-9000系列超纯水系统 美国Milli-Q公司。
1.2 实验方法
1.2.1 菌株活化
将6 株在−80 ℃冷冻保存的芽孢杆菌于室温下进行解冻,用接种环沾取菌液在LB营养琼脂平板上划线,在36±1 ℃培养24 h后将单菌落挑入LB肉汤中,于36±1 ℃培养24 h,完成第一次液体扩培;取200 µL扩培菌液移入至新的LB肉汤,于36±1 ℃培养24 h,完成第二次活化,备用。
1.2.2 基本特性
1.2.2.1 生长曲线测定
取0.2 mL活化菌液接种至5 mL的LB肉汤中,分别放在20 ℃和37 ℃温度条件下培养24 h,每3 h取样测定600 nm下的吸光度。
1.2.2.2 产酸能力测定
取0.5 mL活化后的菌液接种至5 mL的LB肉汤中,放置于20 ℃和37 ℃下培养,在第0、6、12、24、36、48 h取样,8000×g离心5 min,取上清测量pH,结果以不添加菌株的LB肉汤的空白组上清pH数值减去测试菌株的上清pH数值表示。
1.2.2.3 产气能力测定
参考CHEN等[18]的方法并稍作改动,在玻璃试管中注入5 mL的LB肉汤并放入Durham管,接种菌液前保证Durham管内无气泡,取0.8 mL活化后的菌液接种至灭菌后的玻璃试管中,用橡胶塞密封;将所有的玻璃试管置于37 ℃环境中培养,在第12、24、36、48和72 h测量Durham管中的气柱高度。
1.2.3 腐败酶
1.2.3.1 蛋白酶活力测定
液体法测蛋白酶参照国标《GB/T 23527-2009 蛋白酶制剂》中提供的福林法并稍作改良。
活化后的菌液用8500×g离心5 min,上清即为粗酶液。样品组每管加入0.1 mL酪蛋白溶液,空白组每管加入0.2 mL三氯乙酸溶液,于40±0.2 ℃的温度下孵育10 min后用福林溶液显色,待显色完全,在680 nm测定吸光度。制作蛋白酶液体法标准曲线,得出浓度与吸光度之间的拟合公式为Y=0.0053X−0.0024,拟合度R2>0.99,拟合程度较好。根据标准曲线计算出最终的酶活,单位为U/mL。
固体法测蛋白酶活力通过观察菌落在乳粉琼脂平板和熟鸡肉琼脂平板(cooked-chicken juice agar,CJA)上呈现出的分解圈大小,确定蛋白酶活力强弱。乳粉琼脂平板制法参考ELEGBELEYE等[19]方法;CJA平板参考王光宇[20]的生鸡肉琼脂平板(raw-chicken juice agar,RJA)法并改进。
乳粉琼脂平板:分别配制15%脱脂牛奶和LB营养琼脂培养基,分开灭菌后以1:9的比例混匀,倒入无菌培养皿制成乳粉琼脂平板。
CJA:将熟制后的鸡胸肉与0.1 mol/L磷酸盐缓冲液(PBS,pH6.2)以1:40(W/V)的比例匀浆(6000×g,3 min);匀浆液过滤后将滤液与等体积的3%琼脂溶液充分混合,灭菌完成后倒入无菌培养皿制成熟鸡肉琼脂平板。
取活化好的菌液5 µL滴加到乳粉琼脂平板和CJA上,分别放置于20 ℃和37 ℃下培养。每24 h观察记录菌落生长情况和分解圈形成。
1.2.3.2 脂肪酶活力测定
取活化后的菌液,向脂酶细菌微量生化鉴定管注入10 µL菌液,注入菌液时尽量使菌液流过鉴定管内的培养基斜面,充分与指示剂混合后用封口膜密封生化鉴定管。按照试剂盒说明书,将鉴定管置于37 ℃培养48 h,并与空白组对比,观察颜色变化。
1.2.3.3 淀粉酶活力测定
参考DONG等[21]测定芽孢杆菌淀粉酶活力的方法,取活化菌液5 µL滴加到淀粉琼脂平板上,分别放置于20 ℃和37 ℃下培养。每24 h取样,用卢戈氏碘液染色,观察记录菌落生长情况和分解圈形成。
1.3 数据处理
试验数据采用Microsoft office软件进行数据整理,每个指标重复3次,采用平均值±标准差形式表示;分解圈面积采用Image J软件进行计算;用IBM SPSS Statistics 2进行统计分析,采用最小显著差数测验法(LSD)检验不同处理组之间的显著性水平(P<0.05),采用Origin2019b作图。
2. 结果与分析
2.1 基本特性
2.1.1 生长曲线
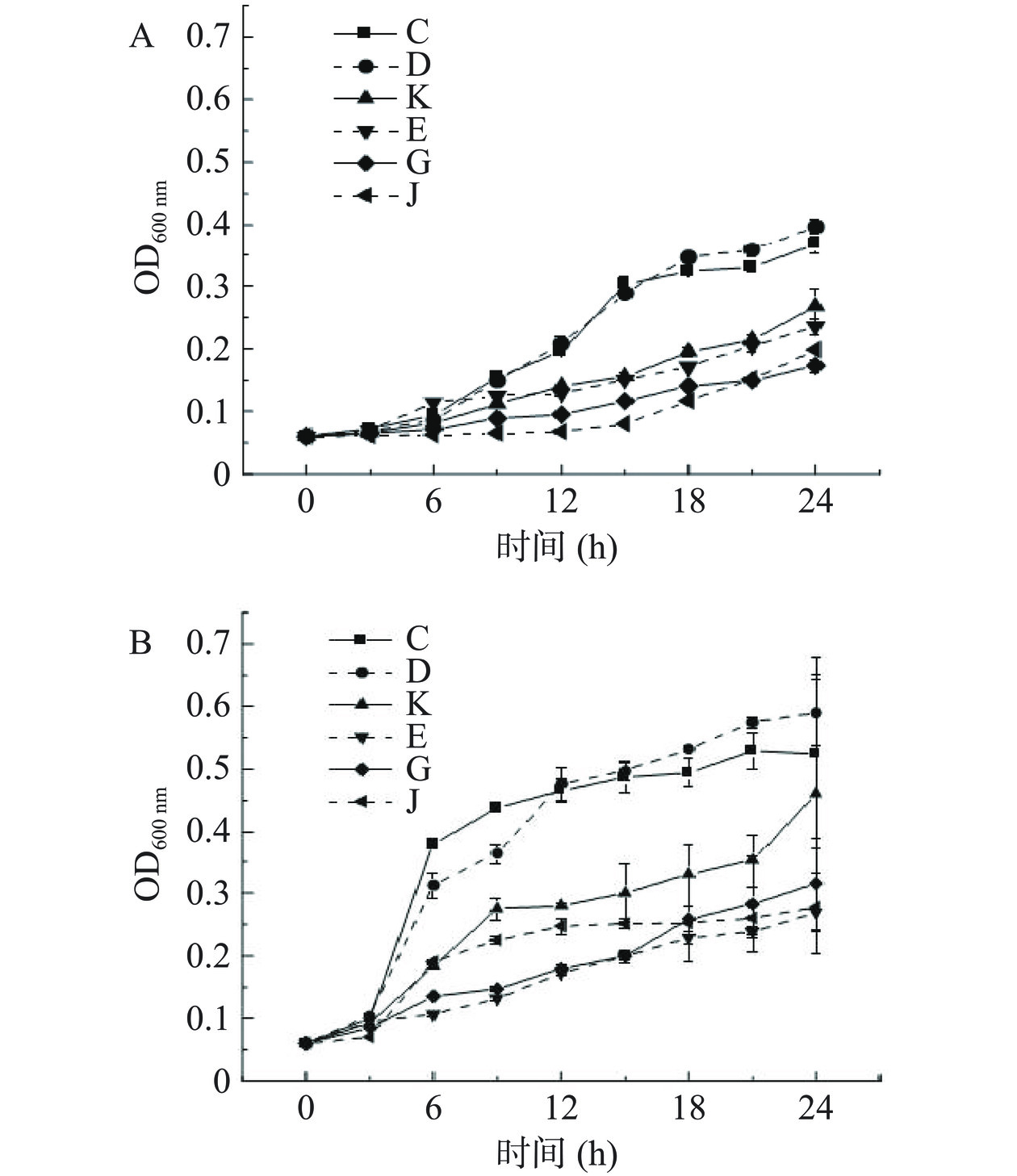
从图1中可以观察到细菌生长周期中的迟缓期、对数期和稳定期,6株芽孢杆菌的生长速率各不相同。在两个培养温度下,蜡样芽孢杆菌(C)和沙福芽孢杆菌(D)生长速率较快,在培养第3 h左右进入对数生长期;而在20 ℃培养温度下,地衣芽孢杆菌(J)生长速率较慢,第12 h左右才进入对数期且OD值较低;在37 ℃培养温度下,沙福芽孢杆菌(E)的生长速率较慢,24 h内生长曲线上升缓慢。
![]() 图 1 20 ℃(A)和37 ℃(B)条件下6株芽孢杆菌的生长曲线(n=3)Figure 1. Growth curves of 6 strains of Bacillus in 20 ℃ (A) and 37 ℃ (B) (n=3)
图 1 20 ℃(A)和37 ℃(B)条件下6株芽孢杆菌的生长曲线(n=3)Figure 1. Growth curves of 6 strains of Bacillus in 20 ℃ (A) and 37 ℃ (B) (n=3)蜡样芽孢杆菌(C)的生长情况与前人研究结果一致[22−23],在37 ℃培养条件下,前2 h处于滞后期,在6 h左右进入稳定期。李宇华[24]在LB培养基中测定吸光度发现,蜡样芽孢杆菌的生长速度比地衣芽孢杆菌更快,说明蜡样芽孢杆菌有更强的生长能力,与本试验结论一致。
2.1.2 产酸
本试验中以相同条件下空白组与菌株pH的差值体现菌株的产酸能力,表2与表3展示了在不同温度下6株菌48 h内的产酸情况。根据显著性分析,在20 ℃下培养48 h,蜡样芽孢杆菌(C)、高山芽孢杆菌(K)、沙福芽孢杆菌(E)和地衣芽孢杆菌(J)的pH变化显著高于其他2株菌(P<0.05),其中蜡样芽孢杆菌(C)和地衣芽孢杆菌(J)pH较空白组下降0.78;在37 ℃组中,培养至48 h的蜡样芽孢杆菌(C)pH下降值仍显著高于其他5株菌(P<0.05),为0.71。根据以上分析,蜡样芽孢杆菌(C)的产酸能力显著强于其他5株菌。
表 2 20 ℃条件下6株菌产酸能力Table 2. Acid production capacity 6 strains of Bacillus in 20 ℃菌株编号 pH下降值(与空白组pH相比) 6 h 12 h 24 h 30 h 36 h 48 h 蜡样芽孢杆菌(C) 0.13±0.02d 0.42±0.03a 0.77±0.03b 0.69±0.05b 0.74±0.04a 0.78±0.03a 沙福芽孢杆菌(D) 0.48±0.03a 0.39±0.02a 0.45±0.03d 0.47±0.04d 0.58±0.02b 0.64±0.05b 高山芽孢杆菌(K) 0.26±0.01b 0.39±0.02a 0.58±0.04c 0.62±0.06c 0.73±0.06a 0.76±0.04a 沙福芽孢杆菌(E) 0.22±0.02c 0.35±0.02b 0.77±0.03b 0.69±0.02b 0.72±0.05a 0.77±0.02a 解淀粉芽孢杆菌(G) 0.49±0.01a 0.35±0.02b 0.42±0.03d 0.48±0.04d 0.47±0.04c 0.51±0.08c 地衣芽孢杆菌(J) 0.10±0.02e 0.10±0.04c 0.90±0.03a 0.86±0.04a 0.73±0.06a 0.78±0.03a 注:同列肩标不同小写字母表示同一培养时间不同菌株pH数据间差异达统计学显著水平(P<0.05),n=5;表3同。 表 3 37 ℃条件下6株菌产酸能力Table 3. Acid production capacity 6 strains of Bacillus in 37 ℃菌株编号 pH下降值(与空白组pH相比) 6 h 12 h 24 h 30 h 36 h 48 h 蜡样芽孢杆菌(C) 0.79±0.03a 0.67±0.04b 0.70±0.02a 0.72±0.03a 0.75±0.04a 0.71±0.03a 沙福芽孢杆菌(D) 0.23±0.01d 0.26±0.03e 0.44±0.07c 0.47±0.05c 0.40±0.05c 0.47±0.04bc 高山芽孢杆菌(K) 0.25±0.03d 0.32±0.03d 0.54±0.05b 0.52±0.03c 0.51±0.02b 0.53±0.05b 沙福芽孢杆菌(E) 0.66±0.01b 0.66±0.05b 0.44±0.08c 0.68±0.05a 0.66±0.10a 0.51±0.20b 解淀粉芽孢杆菌(G) 0.51±0.03c 0.45±0.03c 0.51±0.07bc 0.46±0.06c 0.54±0.03b 0.45±0.05bc 地衣芽孢杆菌(J) 0.80±0.02a 0.74±0.05a 0.65±0.06a 0.60±0.06b 0.56±0.11b 0.36±0.08c 在不同温度下6株芽孢杆菌的产酸能力顺序并不完全一致,但总体来看,全部测试菌株都有较强的产酸能力;6 株菌在生长过程中均会经历先产酸导致pH急剧下降,再回升保持pH在一个相对稳定的范围内的过程。杨福华等[25]用与本试验相同的培养基培养芽孢杆菌,同样发现芽孢杆菌生长过程中培养基溶液的pH下降。田甜等[26]和闫瑞等[27]在研究香肠发酵过程中pH的变化时得出相似的结果,他们分析认为在细菌生长后期出现pH的波动可能是菌株内的蛋白酶分解蛋白质产生了碱性的代谢产物,这些代谢产物调节了培养基内的酸碱值,从而保证细菌生长环境的相对稳定。
2.1.3 产气
如图2所示,所有菌株的Durham管内均无气柱产生;同时根据培养过程的观察,可以确定6 株芽孢杆菌在培养过程中均没有气体产生。
2.2 腐败酶活性
芽孢杆菌产生的蛋白酶会使蛋白质分解,引起食品的腐败变质[28−31]。本试验采取液体法、乳粉琼脂平板法、熟鸡肉琼脂平板法三种不同的方法测定蛋白酶活。三种方法底物各不相同,可综合反映芽孢杆菌对于蛋白的腐败能力。
2.2.1 蛋白酶
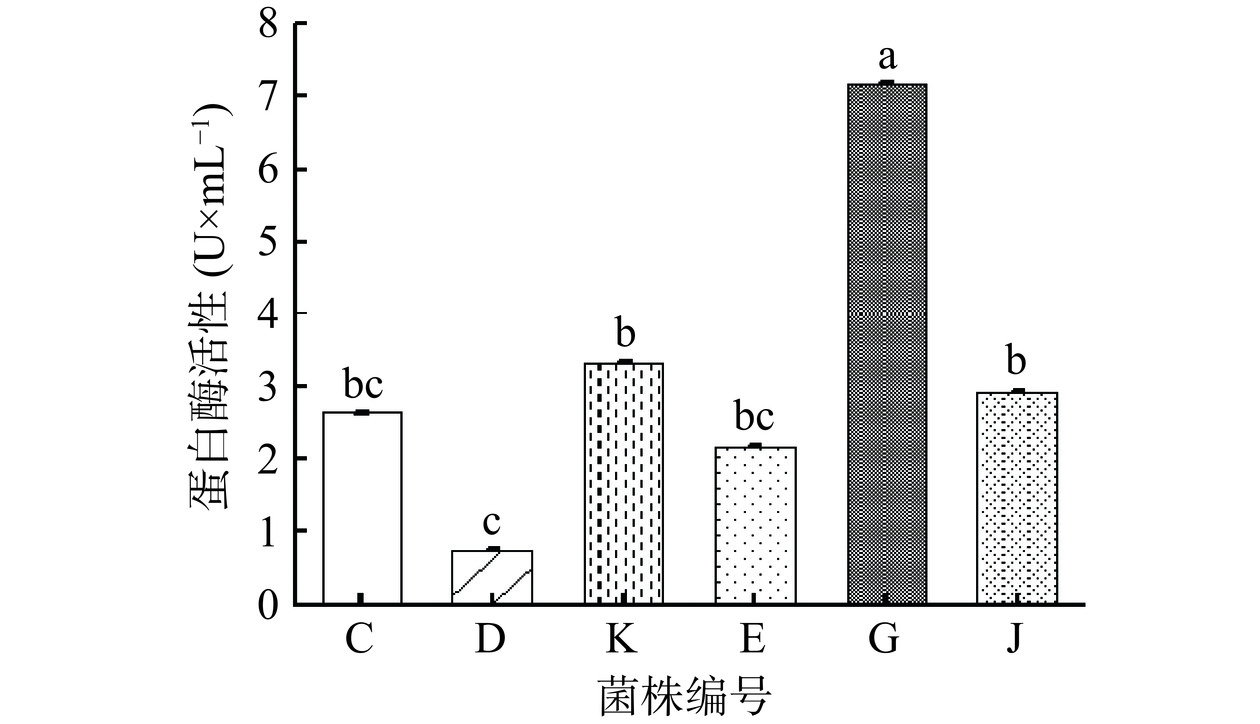
图3展示了6株芽孢杆菌的蛋白酶活力。液体法的反应底物为酪蛋白,通过比较蛋白酶活性数值可以初步判断6株菌的蛋白腐败能力。解淀粉芽孢杆菌(G)表现出的活力最强,为7.17 U/mL,沙福芽孢杆菌(D)的蛋白酶活相比于其他菌较低,为0.75 U/mL,其余4株菌的蛋白酶活在2.16~3.32 U/mL之间。根据试验结果可以初步判定解淀粉芽孢杆菌(G)的蛋白腐败能力最强。然而,液体法只能反映出6株芽孢杆菌对酪蛋白的分解能力。因此,在蛋白酶试验中,需要综合液体法和平板培养的结果,以判断不同菌株之间蛋白酶活力的差异。
![]() 图 3 6 株芽孢杆菌的蛋白酶活性注:菌株蛋白酶活性柱状图中不同字母分别表示显著性差异(P<0.05),n=3。Figure 3. Protease activity of 6 strains of Bacillus
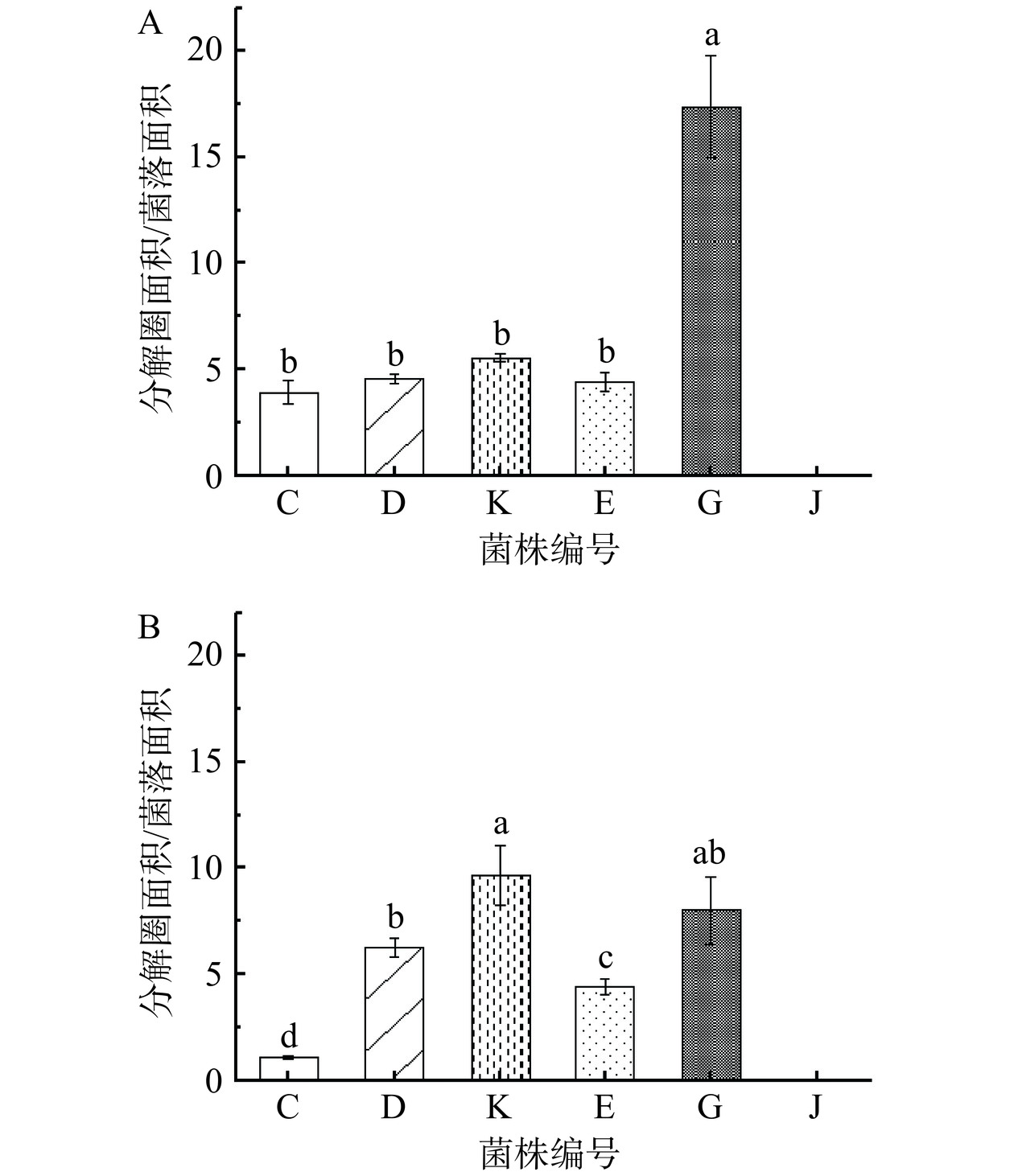
图 3 6 株芽孢杆菌的蛋白酶活性注:菌株蛋白酶活性柱状图中不同字母分别表示显著性差异(P<0.05),n=3。Figure 3. Protease activity of 6 strains of Bacillus图4展示了培养到5 d不同菌株在20 ℃和37 ℃的温度下在乳粉琼脂平板上分解圈面积与菌落面积的比值,以菌株在乳粉琼脂平板上的分解圈直径大小可反映细菌分解蛋白能力的强弱。乳粉琼脂平板法底物为脱脂乳粉,数据分析表明不同分离株的乳粉蛋白分解能力存在显著性差异(P<0.05)。在20 ℃条件下,分解圈面积/菌落面积的比值最大的是解淀粉芽孢杆菌(G),为17.36,分解圈面积/菌落面积的比值由大至小依次为解淀粉芽孢杆菌(G)、高山芽孢杆菌(K)、沙福芽孢杆菌(D)、沙福芽孢杆菌(E)、蜡样芽孢杆菌(C)。在37 ℃条件下,分解圈面积/菌落面积的比值最大的是高山芽孢杆菌(K),为9.64,分解圈面积/菌落面积的比值由大至小依次为高山芽孢杆菌(K)、解淀粉芽孢杆菌(G)、沙福芽孢杆菌(D)、沙福芽孢杆菌(E)、蜡样芽孢杆菌(C)。通过比较分解圈面积与菌落面积的比值大小,可以对6株菌株的蛋白分解能力进行排序。在乳粉琼脂平板试验中,解淀粉芽孢杆菌(G)展现出较强的蛋白酶活性。NGALIMAT 等 [32]在研究中得出了类似结论,发现解淀粉芽孢杆菌的蛋白分解能力较沙福芽孢杆菌和蜡样芽孢杆菌更强,并计算得出解淀粉芽孢杆菌的蛋白酶活力为2.55±0.02 U/mL。
![]() 图 4 20 ℃(A)和37 ℃(B)条件下脱脂乳粉琼脂平板分解圈面积与菌落面积比值注:分解圈面积与菌落面积比值之间的不同字母分别表示显著性差异(P<0.05),n=3。Figure 4. Ratio of decomposition circle area to colony area of skim milk powder agar plate in 20 ℃ (A) and 37 ℃ (B)
图 4 20 ℃(A)和37 ℃(B)条件下脱脂乳粉琼脂平板分解圈面积与菌落面积比值注:分解圈面积与菌落面积比值之间的不同字母分别表示显著性差异(P<0.05),n=3。Figure 4. Ratio of decomposition circle area to colony area of skim milk powder agar plate in 20 ℃ (A) and 37 ℃ (B)CJA的主要营养成分为熟鸡肉匀浆液,可以用于评估菌株对熟鸡肉中蛋白质的腐败的能力。试验所用的6株芽孢杆菌在CJA上的分解圈产生情况如图5所示。结果显示,在适宜的生长温度37 ℃下,经过10 d培养,所有接种至CJA的芽孢杆菌均未产生分解圈。通过计算,本试验中CJA的熟鸡肉添加量为12.5 mg/mL。而鸡肉煮制之后,营养物质会大量流失[33],因此仅以熟鸡胸肉为营养源制备的CJA 平板中可供芽孢杆菌利用的营养物质含量低,仅可维持菌落生长却无明显分解圈产生。
2.2.2 脂肪酶
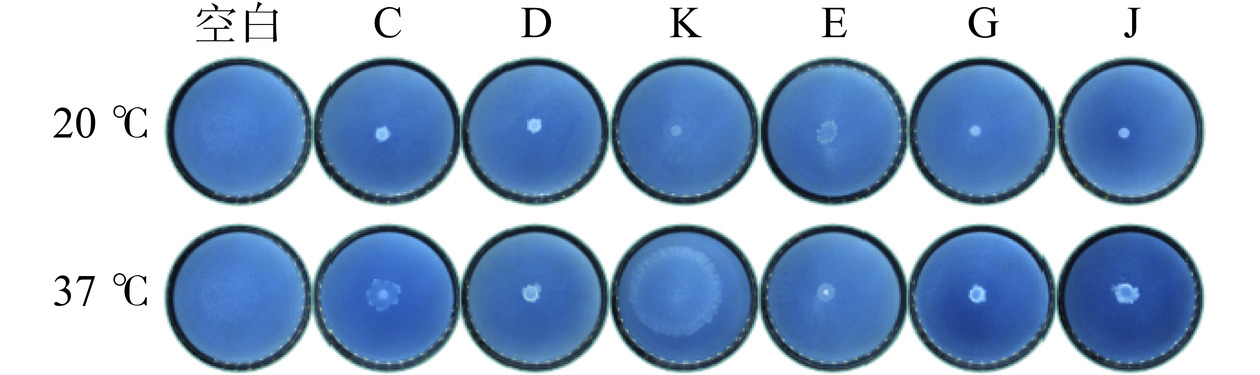
6 株菌株在芽孢杆菌较适宜的生长温度37 ℃条件下培养48 h后全部呈阳性(图6),说明6 株芽孢杆菌可产脂肪酶,具有脂肪腐败能力。芽孢杆菌作为一种常见的胞外脂肪酶来源,部分分离株已被证明具有很强的脂肪酶活力,并用于处理食品工业中富含脂肪的废弃物[33−36]。先前的多项研究[37−40]证明蜡样芽孢杆菌、地衣芽孢杆菌、解淀粉芽孢杆菌和高山芽孢杆菌具有脂肪酶活力,与本试验结论一致。
2.2.3 淀粉酶
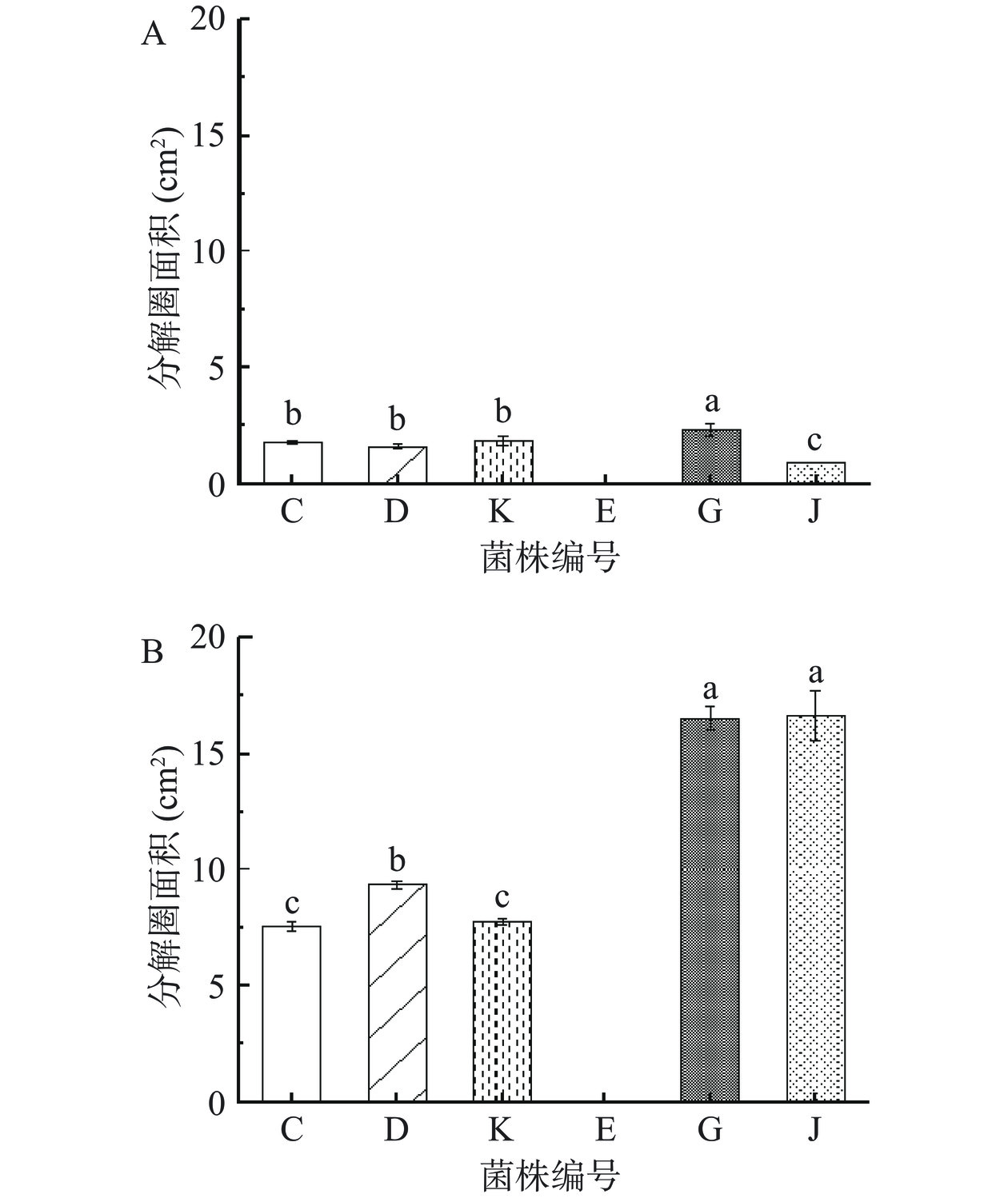
图7展示了在20 ℃和37 ℃条件下培养5 d后,6株芽孢杆菌产生的最大分解圈面积数据。芽孢杆菌作为传统发酵食品中常见的优势微生物[41−42],常见于各种淀粉含量高的食品。SHEN等[43]的研究证明芽孢杆菌具有较强的淀粉分解能力。在本试验的6株芽孢杆菌中,蜡样芽孢杆菌(C)、沙福芽孢杆菌(D)、高山芽孢杆菌(K)、解淀粉芽孢杆菌(G)和地衣芽孢杆菌(J)这5株芽孢杆菌展示出了淀粉分解能力。在20 ℃条件下,解淀粉芽孢杆菌(G)的分解圈面积最大,达到了2.30 cm2,说明在20 ℃的环境温度下,5 株产淀粉酶的菌株中解淀粉芽孢杆菌(G)的淀粉酶活力最强;在37 ℃条件下,解淀粉芽孢杆菌(G)和地衣芽孢杆菌(J)的分解圈面积显著大于另外3株(P<0.05),解淀粉芽孢杆菌(G)的分解圈面积达到了16.53 cm2,地衣芽孢杆菌(J)的分解圈面积达到了16.61 cm2,说明在37 ℃的环境温度下,5 株产淀粉酶的菌株中解淀粉芽孢杆菌(G)、地衣芽孢杆菌(J)的淀粉酶活力最强。
![]() 图 7 20 ℃(A)和37 ℃(B)条件下淀粉琼脂平板分解圈面积注:分解圈面积之间的不同字母分别表示显著性差异(P<0.05),n=3。Figure 7. Decomposition circle area of starch agar plate in 20 ℃ (A) and 37 ℃ (B)
图 7 20 ℃(A)和37 ℃(B)条件下淀粉琼脂平板分解圈面积注:分解圈面积之间的不同字母分别表示显著性差异(P<0.05),n=3。Figure 7. Decomposition circle area of starch agar plate in 20 ℃ (A) and 37 ℃ (B)综合6株菌在两个温度下表现出的淀粉酶活力,6株菌株的淀粉分解能力最强的是解淀粉芽孢杆菌(G)。WANG等[44]通过探究酒曲中不同芽孢杆菌的酶解能力,发现解淀粉芽孢杆菌和地衣芽孢杆菌的葡萄糖淀粉酶活力高于蜡样芽孢杆菌,这与本试验结论一致。LI 等[45]研究指出,解淀粉芽孢杆菌比蜡样芽孢杆菌等拥有更多的胞外淀粉酶和葡萄糖淀粉酶,因此具有更强的淀粉分解能力。
3. 结论
在基本特性试验中,蜡样芽孢杆菌(C)和沙福芽孢杆菌(D)表现出较快的生长速率,在培养的第3 h进入对数生长期。蜡样芽孢杆菌(C)展现了较强的产酸能力,在20 ℃和37 ℃条件下,与空白组相比pH的下降值分别为0.78和0.71;大部分菌株都展现出了蛋白和淀粉腐败能力,其中解淀粉芽孢杆菌(G)的综合腐败能力最强,具体体现为蛋白酶活力为7.17 U/mL,蛋白酶分解圈面积/菌落面积比值和淀粉酶分解圈面积显著高于其他菌株(P<0.05)。
综合试验结果,本研究发现6 株从腐败低温肉制品分离出的芽孢杆菌具有腐败异质性,体现出的生长速率、产酸、蛋白酶活力、淀粉酶活力等均不相同。而不同的理化特性对应不同的防治方法,后续研究中将深入探究芽孢杆菌的腐败机理,并研发更有针对性、更高效的芽孢杆菌控制方法,为提升肉制品安全提供保障。
-
![]()
图 1 20 ℃(A)和37 ℃(B)条件下6株芽孢杆菌的生长曲线(n=3)
Figure 1. Growth curves of 6 strains of Bacillus in 20 ℃ (A) and 37 ℃ (B) (n=3)
![]()
图 3 6 株芽孢杆菌的蛋白酶活性
注:菌株蛋白酶活性柱状图中不同字母分别表示显著性差异(P<0.05),n=3。
Figure 3. Protease activity of 6 strains of Bacillus
![]()
图 4 20 ℃(A)和37 ℃(B)条件下脱脂乳粉琼脂平板分解圈面积与菌落面积比值
注:分解圈面积与菌落面积比值之间的不同字母分别表示显著性差异(P<0.05),n=3。
Figure 4. Ratio of decomposition circle area to colony area of skim milk powder agar plate in 20 ℃ (A) and 37 ℃ (B)
![]()
图 7 20 ℃(A)和37 ℃(B)条件下淀粉琼脂平板分解圈面积
注:分解圈面积之间的不同字母分别表示显著性差异(P<0.05),n=3。
Figure 7. Decomposition circle area of starch agar plate in 20 ℃ (A) and 37 ℃ (B)
表 1 6株芽孢杆菌信息
Table 1 Information of 6 strains of Bacillus
编号 拉丁名 中文名 C Bacillus cereus 蜡样芽孢杆菌 D Bacillus safensis 沙福芽孢杆菌 K Bacillus altitudinis 高山芽孢杆菌 E Bacillus safensis 沙福芽孢杆菌 G Bacillus amyloliquefaciens 解淀粉芽孢杆菌 J Bacillus licheniformis 地衣芽孢杆菌  下载: 导出CSV
下载: 导出CSV
表 2 20 ℃条件下6株菌产酸能力
Table 2 Acid production capacity 6 strains of Bacillus in 20 ℃
菌株编号 pH下降值(与空白组pH相比) 6 h 12 h 24 h 30 h 36 h 48 h 蜡样芽孢杆菌(C) 0.13±0.02d 0.42±0.03a 0.77±0.03b 0.69±0.05b 0.74±0.04a 0.78±0.03a 沙福芽孢杆菌(D) 0.48±0.03a 0.39±0.02a 0.45±0.03d 0.47±0.04d 0.58±0.02b 0.64±0.05b 高山芽孢杆菌(K) 0.26±0.01b 0.39±0.02a 0.58±0.04c 0.62±0.06c 0.73±0.06a 0.76±0.04a 沙福芽孢杆菌(E) 0.22±0.02c 0.35±0.02b 0.77±0.03b 0.69±0.02b 0.72±0.05a 0.77±0.02a 解淀粉芽孢杆菌(G) 0.49±0.01a 0.35±0.02b 0.42±0.03d 0.48±0.04d 0.47±0.04c 0.51±0.08c 地衣芽孢杆菌(J) 0.10±0.02e 0.10±0.04c 0.90±0.03a 0.86±0.04a 0.73±0.06a 0.78±0.03a 注:同列肩标不同小写字母表示同一培养时间不同菌株pH数据间差异达统计学显著水平(P<0.05),n=5;表3同。
下载: 导出CSV
表 3 37 ℃条件下6株菌产酸能力
Table 3 Acid production capacity 6 strains of Bacillus in 37 ℃
菌株编号 pH下降值(与空白组pH相比) 6 h 12 h 24 h 30 h 36 h 48 h 蜡样芽孢杆菌(C) 0.79±0.03a 0.67±0.04b 0.70±0.02a 0.72±0.03a 0.75±0.04a 0.71±0.03a 沙福芽孢杆菌(D) 0.23±0.01d 0.26±0.03e 0.44±0.07c 0.47±0.05c 0.40±0.05c 0.47±0.04bc 高山芽孢杆菌(K) 0.25±0.03d 0.32±0.03d 0.54±0.05b 0.52±0.03c 0.51±0.02b 0.53±0.05b 沙福芽孢杆菌(E) 0.66±0.01b 0.66±0.05b 0.44±0.08c 0.68±0.05a 0.66±0.10a 0.51±0.20b 解淀粉芽孢杆菌(G) 0.51±0.03c 0.45±0.03c 0.51±0.07bc 0.46±0.06c 0.54±0.03b 0.45±0.05bc 地衣芽孢杆菌(J) 0.80±0.02a 0.74±0.05a 0.65±0.06a 0.60±0.06b 0.56±0.11b 0.36±0.08c
下载: 导出CSV
-
[1] 中华人民共和国国家统计局. 中国统计年鉴[M]. 北京:中国统计出版社, 2022. [National Bureau of Statistics of China. China statistical yearbook[M]. Beijing:China Statistics Press, 2022.] National Bureau of Statistics of China. China statistical yearbook[M]. Beijing: China Statistics Press, 2022.
[2] AHMAD M I, FAROOQ S, ALHAMOUD Y, et al. A review on mycoprotein:History, nutritional composition, production methods, and health benefits[J]. Trends in Food Science & Technology,2022,121(Mar.):14−29.
[3] 陈坚. 中国食品科技:从2020到2035[J]. 中国食品学报,2019,19(12):1−5. [CHEN Jian. Food science and technology in China:From 2020 to 2035[J]. Journal of Chinese Institute of Food Science and Technology,2019,19(12):1−5.] CHEN Jian. Food science and technology in China: From 2020 to 2035[J]. Journal of Chinese Institute of Food Science and Technology, 2019, 19(12): 1−5.
[4] SANLIER N, SORMAZ Ü, GÜNES E. The effect of food safety education on food safety knowledge, attitudes, behaviors of individuals who work in food and beverage departments in Turkey[J]. International Journal of Gastronomy and Food Science,2020,22:100259. doi: 10.1016/j.ijgfs.2020.100259
[5] GODFRAY H C J, AVEYARD P, GARNETT T, et al. Meat consumption, health, and the environment[J]. Science,2022,361(Jul. 20 TN. 6399):243.
[6] 贾瑞娟. 山西老陈醋源芽孢菌抑菌机理及对腐败菌群体感应抑制的研究[D]. 太原:山西农业大学, 2019. [JIA Ruijuan. Antibacterial mechanism of Bacillus from Shanxi aged vinegar and its inhibition quorum sensing of spoilage bacteria[D]. Taiyuan:Shanxi Agricultural University, 2019.] JIA Ruijuan. Antibacterial mechanism of Bacillus from Shanxi aged vinegar and its inhibition quorum sensing of spoilage bacteria[D]. Taiyuan: Shanxi Agricultural University, 2019.
[7] MARKLAND S, HOOVER D. The diverse faces of Bacillus cereus[M]. Elsevier Inc.,2016:45−59.
[8] 李冉, 朱和源, 叶可萍, 等. 气调包装狮子头冷藏过程中微生物变化及菌群结构分析[J]. 食品工业科技,2021,42(11):99−105. [LI Ran, ZHU Heyuan, YE Keping, et al. Analysis of microbial counts and bacterial community in modified atmosphere packaging meatballs[J]. Science and Technology of Food Industry,2021,42(11):99−105.] LI Ran, ZHU Heyuan, YE Keping, et al. Analysis of microbial counts and bacterial community in modified atmosphere packaging meatballs[J]. Science and Technology of Food Industry, 2021, 42(11): 99−105.
[9] 甯雨荞, 田靓, 袁先铃, 等. 市售笋子烧牛肉中优势腐败菌的分离与鉴定[J]. 农产品加工,2021(8):51−55. [NING Yuqiao, TIAN Liang, YUAN Xianling, et al. Isolation and identification of dominant spoilage bacteria in commercially available roasted beef with bamboo shoots[J]. Farm Products Processing,2021(8):51−55.] NING Yuqiao, TIAN Liang, YUAN Xianling, et al. Isolation and identification of dominant spoilage bacteria in commercially available roasted beef with bamboo shoots[J]. Farm Products Processing, 2021(8): 51−55.
[10] MARKLAND S M, HOOVER D G. Chapter 4-Bacillus cereus mechanisms of resistance to food processing[M]//SAVINI V. The diverse faces of Bacillus cereus. Academic Press,2016:45-59.
[11] 徐靖, 牛邦彦, 张亚南, 等. 芽孢杆菌属Bacillus分类学研究进展[J]. 中国土壤与肥料,2022(12):225−237. [XU Jing, NIU Bangyan, ZHANG Yanan, et al. Advances in taxonomy of genus Bacillus[J]. Soil and Fertilizer Sciences in China,2022(12):225−237.] doi: 10.11838/sfsc.1673-6257.22079 XU Jing, NIU Bangyan, ZHANG Yanan, et al. Advances in taxonomy of genus Bacillus[J]. Soil and Fertilizer Sciences in China, 2022(12): 225−237. doi: 10.11838/sfsc.1673-6257.22079
[12] KAUR C, VELMURUGAN N, KUMAR S, et al. Bacillus aequororis sp. nov. isolated from marine sediment[J]. Current Microbiology:An International Journal,2014,69(5):758−762.
[13] 全拓. 肉制品中主要微生物的检测与研究[D]. 重庆:西南大学, 2012. [QUAN Tuo. The detection and research of main microorganisms in meat products[D]. Chongqing:Southwest University, 2012.] QUAN Tuo. The detection and research of main microorganisms in meat products[D]. Chongqing: Southwest University, 2012.
[14] 王茜. 即食小龙虾软包装体系热杀菌工艺优化[D]. 无锡:江南大学, 2022. [WANG Qian. Thermal sterilization technology optimization of instant crayfish flexible packaging system[D]. Wuxi:Jiangnan University, 2022.] WANG Qian. Thermal sterilization technology optimization of instant crayfish flexible packaging system[D]. Wuxi: Jiangnan University, 2022.
[15] WEBB M D, BARKER G C, GOODBURN K E, et al. Risk presented to minimally processed chilled foods by psychrotrophic Bacillus cereus[J]. Trends in Food Science & Technology,2019,93:94−105.
[16] 杨啸吟, 张一敏, 梁荣蓉, 等. 包装冷却肉中微生物腐败及其挥发性气味的研究进展[J]. 食品科学,2021,42(1):285−293. [YANG Xiaoyin, ZHANG Yimin, LIANG Rongrong, et al. Microbial spoilage and formation and detection of volatile odor compounds in packaged chilled meat:A review[J]. Food Science,2021,42(1):285−293.] doi: 10.7506/spkx1002-6630-20200207-046 YANG Xiaoyin, ZHANG Yimin, LIANG Rongrong, et al. Microbial spoilage and formation and detection of volatile odor compounds in packaged chilled meat: A review[J]. Food Science, 2021, 42(1): 285−293. doi: 10.7506/spkx1002-6630-20200207-046
[17] SHAO Liangting, CHEN Shanshan, WANG Haodong, et al. Advances in understanding the predominance, phenotypes, and mechanisms of bacteria related to meat spoilage[J]. Trends in Food Science & Technology,2021(Pt.B):822−832.
[18] CHEN Shanshan, LIU Silu, MA Jiaming, et al. Evaluation of the spoilage heterogeneity of meat-borne Leuconostoc mesenteroides by metabonomics and in-situ analysis[J]. Food Research International,2022,156(Jun.):111365.1−111365.9.
[19] ELEGBELEYE J A, BUYS E M. Potential spoilage of extended shelf-life (ESL) milk by Bacillus subtilis and Bacillus velezensis[J]. LWT,2022,153(Jan.):112487.
[20] 王光宇. 气调包装对冷鲜鸡肉中莓实假单胞菌致腐效应的抑制机制[D]. 南京:南京农业大学, 2018:28−29. [WANG Guangyu. Inhibition mechanisms of MAP against chilled chicken spoilage associated with Pseudomonas fragi[D]. Nanjing:Nanjing Agricultural University, 2018:28−29.] WANG Guangyu. Inhibition mechanisms of MAP against chilled chicken spoilage associated with Pseudomonas fragi[D]. Nanjing: Nanjing Agricultural University, 2018: 28−29.
[21] DONG Honghao, ZHU Hong, HE Hailun, et al. Identification and evaluation of spoilage potential of four Bacillus strains isolated from slimy rice noodles[J]. Food Microbiology,2023,110:104160. doi: 10.1016/j.fm.2022.104160
[22] DIAO Mingming, QI Danping, XU Miaomiao, et al. Antibacterial activity and mechanism of monolauroyl-galactosylglycerol against Bacillus cereus[J]. Food Control,2018,85:339−344. doi: 10.1016/j.foodcont.2017.10.019
[23] SHU Q, NIU Y, ZHAO W, et al. Antibacterial activity and mannosylerythritol lipids against vegetative cells and spores of Bacillus cereus[J]. Food Control,2019,106:106711. doi: 10.1016/j.foodcont.2019.106711
[24] 李宇华. 葡萄糖对芽孢杆菌属内互作影响的研究[D]. 咸阳:西北农林科技大学, 2022:15−16. [LI Yuhua. Effect of glucose on the interspecific interaction within Bacillus genus[D]. Xianyang:Northwest A&F University, 2022:15−16.] LI Yuhua. Effect of glucose on the interspecific interaction within Bacillus genus[D]. Xianyang: Northwest A&F University, 2022: 15−16.
[25] 杨福华, 刘韶娜, 廖隽锐, 等. 芽孢杆菌 YF-15 的筛选, 鉴定及对蚕豆秸秆青贮营养品质的影响[J]. 饲料研究,2023,46(9):121−125. [YANG Fuhua, LIU Shaona, LIAO Junrui, et al. Screening and identification of Bacillus YF-15 and its effect on nutritional quality of faba bean straw[J]. Feed Research,2023,46(9):121−125.] YANG Fuhua, LIU Shaona, LIAO Junrui, et al. Screening and identification of Bacillus YF-15 and its effect on nutritional quality of faba bean straw[J]. Feed Research, 2023, 46(9): 121−125.
[26] 田甜, 张雅琳, 潘攀, 等. 微生物复合发酵剂对香肠的食用安全性分析[J]. 食品工业,2020,41(6):329−333. [TIAN Tian, ZHANG Yalin, PAN Pan, et al. Analysis of food safety of compound fermenting agent on sausage[J]. The Food Industry,2020,41(6):329−333.] TIAN Tian, ZHANG Yalin, PAN Pan, et al. Analysis of food safety of compound fermenting agent on sausage[J]. The Food Industry, 2020, 41(6): 329−333.
[27] 闫瑞, 唐敏, 陈东方, 等. 白萝卜粉作为亚硝酸盐来源制备猪肉发酵干香肠[J]. 食品科学,2023,44(8):101−108. [YAN Rui, TANG Min, CHEN Dongfang, et al. Preparation of dry fermented pork sausage using radish powder as a source of nitrite[J]. Food Science,2023,44(8):101−108.] doi: 10.7506/spkx1002-6630-20220713-138 YAN Rui, TANG Min, CHEN Dongfang, et al. Preparation of dry fermented pork sausage using radish powder as a source of nitrite[J]. Food Science, 2023, 44(8): 101−108. doi: 10.7506/spkx1002-6630-20220713-138
[28] FEI Peng, XU Yunfeng, ZHAO Shengjuan, et al. Olive oil polyphenol extract inhibits vegetative cells of Bacillus cereus isolated from raw milk[J]. Journal of Dairy Science,2019,102(5):3894−3902. doi: 10.3168/jds.2018-15184
[29] MOSCHONAS G, LIANOU A, NYCHAS G-J E, et al. Spoilage potential of Bacillus subtilis in a neutral-pH dairy dessert[J]. Food Microbiology,2021,95:103715. doi: 10.1016/j.fm.2020.103715
[30] ZHAI Zhengyuan, CUI Chang, LI Xueli, et al. Prevalence, antimicrobial susceptibility, and antibiotic resistance gene transfer of Bacillus strains isolated from pasteurized milk[J]. Journal of Dairy Science,2023,106(1):75−83. doi: 10.3168/jds.2022-22199
[31] WANG Jing, SU Yujie, GU Luping, et al. The inhibition of cell-free supernatants of several lactic acid bacteria on the selected psychrophilic spoilage bacteria in liquid whole egg[J]. Food Control,2020,123(5):107753.
[32] NGALIMAT M S, RAHMAN R N Z R A, YUSOF M T, et al. Characterisation of bacteria isolated from the stingless bee, Heterotrigona itama, honey, bee bread and propolis[J]. PeerJ,2019,7(12):e7478.
[33] JAYASENA D D, JUNG S, KIM S H, et al. Endogenous functional compounds in Korean native chicken meat are dependent on sex, thermal processing and meat cut[J]. Journal of the Science of Food & Agriculture,2015,95(4):771−775.
[34] SARASWAT R, BHUSHAN I, GUPTA P, et al. Production and purification of an alkaline lipase from Bacillus sp. for enantioselective resolution of (±)-Ketoprofen butyl ester[J]. 3 Biotech,2018,8(12):491−491. doi: 10.1007/s13205-018-1506-6
[35] BALAJI L, CHITTOOR J T, JAYARAMAN G. Optimization of extracellular lipase production by halotolerant Bacillus sp. VITL8 using factorial design and applicability of enzyme in pretreatment of food industry effluents[J]. Preparative Biochemistry & Biotechnology:An International Journal for Rapid Communication,2020,50(7):708−716.
[36] SINGH P, PATEL V, SHAH V, et al. A solvent-tolerant alkaline lipase from Bacillus sp. DM9K3 and its potential applications in esterification and polymer degradation[J]. Applied Biochemistry and Microbiology,2019,55(6):603−614.
[37] DEMIRKAN E, ÇETINKAYA A A, ABDOU M. Lipase from new isolate Bacillus cereus ATA179:Optimization of production conditions, partial purification, characterization and its potential in the detergent industry[J]. Turkish Journal of Biology,2021,45(3):287−300. doi: 10.3906/biy-2101-22
[38] MAZHAR H, ULLAH I, ALI U, et al. Optimization of low-cost solid-state fermentation media for the production of thermostable lipases using agro-industrial residues as substrate in culture of Bacillus amyloliquefaciens[J]. Biocatalysis and Agricultural Biotechnology,2023,47:102559. doi: 10.1016/j.bcab.2022.102559
[39] ZHAO Junxin, LIU Shichang, GAO Yifang, et al. Characterization of a novel lipase from Bacillus licheniformis NCU CS-5 for applications in detergent industry and biodegradation of 2, 4-D butyl ester[J]. International Journal of Biological Macromolecules:Structure, Function and Interactions,2021,176:126−136.
[40] ESAKKIRAJ P, USHA R, PALAVESAM A, et al. Solid-state production of esterase using fish processing wastes by Bacillus altitudinis AP-MSU[J]. Food and Bioproducts Processing,2012,90(3):370−376. doi: 10.1016/j.fbp.2011.12.008
[41] 吴博华, 蒋雪薇, 张金玉, 等. 促酱醪发酵芽孢杆菌的筛选及应用[J]. 食品科学,2021,42(6):134−141. [WU Bohua, JIANG Xuewei, ZHANG Jinyu, et al. Screening and application of Bacillus spp. promoting moromi fermentation[J]. Food Science,2021,42(6):134−141.] doi: 10.7506/spkx1002-6630-20200106-076 WU Bohua, JIANG Xuewei, ZHANG Jinyu, et al. Screening and application of Bacillus spp. promoting moromi fermentation[J]. Food Science, 2021, 42(6): 134−141. doi: 10.7506/spkx1002-6630-20200106-076
[42] 向慧平, 林宜锦, 关统伟, 等. 四川浓香型大曲生产中酵母菌、芽孢杆菌与工艺指标的关联性分析[J]. 食品科学,2020,41(2):196−201. [XIANG Huiping, LIN Yijin, GUAN Tongwei, et al. Diversities of culturable yeast and Bacillus and their relationship with process parameters during the production of Sichuan Luzhou-flavor Daqu[J]. Food Science,2020,41(2):196−201.] doi: 10.7506/spkx1002-6630-20181217-179 XIANG Huiping, LIN Yijin, GUAN Tongwei, et al. Diversities of culturable yeast and Bacillus and their relationship with process parameters during the production of Sichuan Luzhou-flavor Daqu[J]. Food Science, 2020, 41(2): 196−201. doi: 10.7506/spkx1002-6630-20181217-179
[43] SHEN Guanghui, ZHENG Lijun, LI Shanshan, et al. The role of soy protein degradation caused by spoilage Bacillus amyloliquefaciens in texture deterioration of Yuba, a soy product[J]. LWT,2020,123:109108. doi: 10.1016/j.lwt.2020.109108
[44] WANG Zongmin, WANG Chengtao, SHEN Caihong, et al. Microbiota stratification and succession of amylase-producing Bacillus in traditional Chinese Jiuqu (fermentation starters)[J]. Journal of the Science of Food and Agriculture,2020,100(8):3544−3553. doi: 10.1002/jsfa.10405
[45] LI Zhen, SIEPMANN F B, TOVAR L E R, et al. Effect of copy number of the spoVA2mob operon, sourdough and reutericyclin on ropy bread spoilage caused by Bacillus spp.[J]. Food Microbiology,2020,91:103507. doi: 10.1016/j.fm.2020.103507
 下载:
下载:









计量
- 文章访问数: 0
- HTML全文浏览量: 0
- PDF下载量: 0
