


Efficient Soluble Expression and Fermentation Conditions of D-Allulose 3-Epimerase in Escherichia coli
-
摘要: 将来源于Clostridium bolteae ATCCBAA-613的DAEase基因序列经密码子优化合成,以pCold TF为表达载体,冷休克启动子CspA低温诱导DAEase基因在大肠杆菌(Escherichia coli)BL21 (DE3)中表达,得到高效可溶性的重组Cb-DAEase并利用镍柱亲和层析分离纯化。结果表明,Cb-DAEase最适pH和温度为7.0和55 ℃,Co2+能够显著(P<0.05)增强酶活力。对培养条件进行优化得到,在7 g/L甘油、10 g/L酵母膏、1%接种量、0.25 mmol/LIPTG、诱导前培养5 h的条件下,Cb-DAEase活力达到(10.11±0.02)U/g,比优化前(1.38±0.01) U/g提高了7.33倍;以120 g/L的D-果糖为底物全细胞催化0.5 h后,D-阿洛酮糖产量为(11.47±0.04)g/L,比优化前(1.03±0.02)g/L提高了11.14倍。基于冷休克表达策略构建的重组菌经发酵优化后Cb-DAEase活力显著(P<0.05)提高,为高效制备D-阿洛酮糖提供了理论支持。Abstract: The DAEase gene sequence derived from Clostridium bolteae ATCCBAA-613 was synthesized by codon optimization. Using pCold TF as the expression vector, the cold-shock promoter CspA induced the expression of the DAEase gene in Escherichia coli BL21(DE3) at low temperature. Then, the highly soluble recombinant Cb-DAEase was obtained and purified by Ni-chelating affinity chromatography. Results showed that, the Cb-DAEase exhibited maximum activity at pH7.0 and 55 ℃. Additionally, the Cb-DAEase showed different sensitivities to the various metal ions when Co2+ was able to significantly (P<0.05) enhance its enzyme activity. The optimum fermentation conditions were determined as follows, 7 g/L glycerol, 10 g/L yeast extract, 1% inoculation volume, 0.25 mmol/L IPTG inducer, and incubation 5 h before the induction. Eventually, the secretion level of Cb-DAEase reached (10.11±0.02) U/g, which was 7.33-fold higher than that control (1.38±0.01) U/g. Through optimizing conditions, D-allulose was produced effectively by the whole-cell biotransformation system when 120 g/L D-fructose was used as the substrate for 0.5 h, the yield reached (11.47±0.04) g/L, which was 11.14-fold higher than that control (1.03±0.02) g/L. Overall, the recombinant strain constructed based on cold-shock expression increased significantly (P<0.05) Cb-DAEase activity after fermentation optimization, which would provide a theoretical basis for the efficient preparation of D-allulose.
-
近年来,D-阿洛酮糖由于其多种生理功能而受到越来越多的关注,可作为抗癌剂、有效免疫抑制剂以及抗氧化剂,市场潜力巨大,目前针对化工合成法纯化复杂、伴有副产物产生的缺点,生物合成法生产D-阿洛酮糖具有良好的应用前景[1]。D-阿洛酮糖3-差向异构酶(D-allulose 3-epimerase,DAEase)是催化D-果糖和D-阿洛酮糖进行可逆反应的异构化酶[1]。近年来,陆续有不同来源DAEase在不同宿主中表达的报道[2-4]。其中,Wei等[2]在枯草芽孢杆菌中成功表达了来自Clostridium bolteae 和Dorea sp.的两个DAEase基因,并通过启动子强度优化将DAEase的产量提高约20%。然而在枯草芽孢杆菌表达系统中,重组质粒具有不稳定性,在大规模生产中容易丢失或发生结构改变,不利于相关的基因表达和调控研究[5]。酿酒酵母作为食品级微生物,Juneja等[4]在酿酒酵母中成功表达了DAEase,在55 ℃时将D-果糖转化为D-阿洛酮糖的转化率为26.6%。然而在酵母表达系统中,外源蛋白易于糖基化及发酵过程中产生乙醇,并会导致目标蛋白产量下降[5]。
与其它宿主相比,大肠杆菌具有外源基因表达效率高、蛋白分泌量大和基因操作简单等优点[5-6]。Zhang等[7]使用重组大肠杆菌表达来源于Dorea sp. CAG317的DAEase,在pH6.0的酸性条件下酶活性最高,达到了803 U/mg;Park等[8]在重组大肠杆菌中成功表达了来自Agrobacterium tumefaciens的DAEase双位点突变体(I33L/S213C),培养24 h后重组大肠杆菌中DAEase和粗蛋白的表达量分别为8.6%和37.0%(w/w)。目前大肠杆菌表达系统主要存在的问题是蛋白易发生错误折叠而形成不溶性包涵体,导致产量偏低[9]。将包涵体通过树脂和凝胶色谱分离纯化耗时耗力,且复性过程易造成肽链间的错误折叠[10];此外,蛋白质的正确折叠需在基因表达速率和蛋白溶解度之间达到平衡,基因操作难度大[11]。尽管Tseng等[12]将α-突触核蛋白酸性尾部衍生的肽段与Agrobacterium sp. ATCC31749的DAEase融合,其在大肠杆菌中的可溶性表达增加了1.57倍,但酶的分离纯化步骤却较为繁琐,不利于生产应用。
据报道,温度会影响蛋白质的合成速率、折叠动力学和蛋白质降解的疏水作用[13],低温会为蛋白质折叠提供充足时间[14]。闫真等[15]发现,在17 ℃下诱导表达的重组酶比活明显高于37 ℃下诱导表达的比活,说明低温有利于提高可溶性蛋白的产量。低温可以降低包涵体的数量,然而低温下菌体生长缓慢会导致单位时间的表达水平降低[16]。CspA是37 ℃时处于抑制状态的冷休克蛋白,在低温(15 ℃)下会被激活诱导重组蛋白表达[17]。包含CspA启动子的pCold-TF是一种融合冷休克表达载体,目标基因被冷激诱导表达之前,细菌可在37 ℃快速生长;另外,TF分子伴侣可以作为可溶性融合标签表达,防止蛋白质在密集细胞环境中发生错误折叠和聚集,与蛋白质结合从而使其稳定折叠,并在发挥作用后自行离开目的蛋白,有利于后续的分离纯化[17]。因此,为了提高DAEase的产量,本文选择冷休克表达质粒pCold TF作为表达载体,以实现DAEase在大肠杆菌表达系统内的高效可溶性表达,并对全细胞催化D-果糖生产D-阿洛酮糖的条件进行优化,为其高效工业化生产提供理论依据。
1. 材料与方法
1.1 材料与仪器
T4 DNA连接酶、SDS-PAGE变性丙烯酰胺凝胶快速制备试剂盒、5×蛋白加样缓冲液、Real Band 蛋白预染Marker、异丙基-β-D-硫代半乳糖苷(IPTG)、氨苄青霉素(Amp) 生工生物工程(上海)股份有限公司(Sangon Biotech);限制性核酸内切酶EcoR Ⅰ、Hind Ⅲ TaKaRa公司;Ni Seoharose 6 Fsat Flow蛋白纯化树脂 美国GE公司;D-阿洛酮糖标品(99%) 色谱纯 Sigma公司;其他试剂 均为国产分析纯;大肠杆菌Escherichia coli DH5α、BL21 (DE3)感受态细胞、冷休克表达载体pCold TF 由本实验室保存;引物合成和核酸测序 由生工生物工程(上海)股份有限公司完成;LB培养基:10 g/L 蛋白胨、10 g/L 氯化钠和5 g/L 酵母提取物,pH7.2,121 ℃灭菌20 min,用于大肠杆菌的培养。
GR85DP型高压灭菌锅 智微(厦门)仪器有限公司;Power PacTM Basic型蛋白电泳仪 美国伯乐(BIO-RAD)公司;T100 Thermal Cycler型PCR仪 美国伯乐(BIO-RAD)公司;Waters 2595型高效液相色谱仪 美国沃特世(Waters)公司;722N可见分光光度计 上海仪电分析仪器有限公司。
1.2 实验方法
1.2.1 重组质粒的构建
在NCBI数据库上筛选得到来源于C. bolteae ATCCBAA-613的DAEase基因序列,对其进行了针对E. coli中表达的密码子的优化[18],在C端加上6×His标签,同时在基因序列的5'端与3'端分别引入EcoR Ⅰ和Hind Ⅲ 酶切位点,将优化后的基因序列交由上海生工生物公司合成,连接至载体PUC57上。将优化合成的DAEase基因用EcoR Ⅰ和Hind Ⅲ双酶切,并与经同样酶切的pCold TF载体在T4连接酶的介导下连接,将其转化至E. coli DH5α感受态细胞中,37 ℃培养过夜[19]。次日挑取单克隆于LB培养基(含100 µg/mL Amp)中,37 ℃、200 r/min培养12 h,提取质粒,EcoR Ⅰ和Hind Ⅲ双酶切鉴定目的基因的插入,并送测序。经测序验证后,将构建正确的重组质粒命名为pCold TF-Cb-DAE。
1.2.2 pCold TF-Cb-DAE重组质粒诱导表达
将重组质粒pCold TF-Cb-DAE转入到大肠杆菌E. coli BL21 (DE3)中,37 ℃过夜培养后,取1%的种子液接种于50 mL LB培养基(含100 µg/mL Amp),37 ℃下200 r/min振荡培养至生物量OD600约为0.6~0.8,加入0.25 mmol/L IPTG,15 ℃下200 r/min低温诱导培养,24 h后收集菌液离心,用15 mL 1× PBS缓冲液悬浮菌体,经超声波细胞破壁后的上清液及沉淀经十二烷基硫酸钠-聚丙烯酰胺凝胶电泳(sodium dodecyl sulfate-polyacrylamide gel electrophoresis, SDS-PAGE)鉴定,分析菌体中目的蛋白的表达情况。
1.2.3 Cb-DAEase的纯化
重组菌株经摇瓶培养24 h后,收集上清液用于蛋白质纯化。上清液中存在大量的杂蛋白质,使用Ni Seoharose 6 Fast Flow蛋白纯化树脂对其进行纯化,可以得到高纯度的目的蛋白[20]。采用梯度洗脱法(咪唑浓度20、50、500 mmol/L),低浓度咪唑洗脱液洗脱杂蛋白,高浓度咪唑洗脱液洗脱目的蛋白,SDS-PAGE检测纯化效果[21]。
1.2.4 Cb-DAEase的酶学性质
为进一步验证目的蛋白是否可以催化合成D-阿洛酮糖,以50 g/L D-果糖为底物测酶活力,加入适量纯化的Cb-DAEase和1 mmol/L的Co2+,在55 ℃和pH 7.0条件下催化10 min后,立即沸水浴10 min以终止反应,反应终止液用0.22 µm滤膜过滤后,通过高效液相色谱(high performance liquid chromatography,HPLC)检测D-阿洛酮糖的合成,酶活定义为:每分钟产生1 µmol的D-阿洛酮糖所需的酶量是1 U。检测条件:Waters 2595型液相色谱仪,Sugar-Pak1糖柱,Waters 2414示差折光检测器,85 ℃柱温,流动相为超纯水,流速为0.4 mL/min[22]。
1.2.4.1 最适温度和pH的测定
在pH8.0条件下,测定不同温度(40、45、50、55、60、70、75及80 ℃)条件下的酶活力;在测得的最适反应温度条件下,测定不同pH(4.5、5.0、5.5、6.0、6.5、7.0、7.5、8.0和8.5)条件下的酶活力,将最高酶活力定义为相对酶活100%,其他处理组酶活力/最高酶活力×100%为相对酶活力。
1.2.4.2 金属离子对DAEase活力的影响
保持上述最适催化温度和pH不变,在标准测定体系中分别加入1 mmol/L不同金属离子(Co2+、Mn2+、Mg2+、Cu2+、Ca2+、Ba2+和Zn2+),催化反应10 min,使用HPLC测定酶活力。以缓冲溶液代替金属离子为对照组,将对照组的酶活力设为相对酶活100%。
1.2.5 重组菌株发酵条件优化
1.2.5.1 碳源对细胞生长和Cb-DAEase活力的影响
在LB培养基的基础上,分别以5.0 g/L的葡萄糖、果糖、乳糖、蔗糖、可溶性淀粉和甘油为碳源,按1%的接种量转接到50 mL培养基(含Amp浓度100 mg/L)中,于37 ℃下200 r/min恒温振荡培养5 h,当OD600为0.6~0.8时加入0.25 mol/L IPTG诱导剂,继续置于15 ℃的摇床中以200 r/min的转速诱导培养24 h,考察不同种类的碳源对酶活力和菌体生长量的影响。酶活力测定同1.2.4节,菌体生长量的测定:将发酵液样品稀释至适宜浓度,使用分光光度计记录OD600,其中稀释倍数以OD600落在0.2~0.8确定。
1.2.5.2 氮源对细胞生长和Cb-DAEase活力的影响
在LB培养基的基础上,以7 g/L的甘油为碳源,分别以10 g/L的酵母膏、牛肉膏、蛋白胨、胰蛋白胨、酪蛋白胨、氯化铵作为氮源,考察不同种类的氮源对酶活力和菌体生长量的影响,其他培养条件同1.2.5.1节。
1.2.5.3 培养条件对Cb-DAEase活力的影响
将培养过夜的重组菌株BL21/TF-Cb接种到200 mL含有100 µg/mL Amp的LB液体培养基中,37 ℃下200 r/min培养至OD600值为0.6~0.8,使用单因素实验考察接种量、IPTG诱导剂浓度和诱导前培养时间等因素对酶活力的影响,将最高酶活力定义为相对酶活100%。在培养基中接种不同量(0.75%、1.0%、1.25%、1.5%、1.75%和2.0%)的种子液,添加0.25 mmol/L IPTG诱导剂,于15 ℃下200 r/min诱导24 h,收集发酵液测定酶活力和菌体生长量;在培养基接种1%的种子液,添加不同浓度(0.1、0.15、0.2、0.25、0.3、0.35及0.4 mmol/L)的IPTG,于15 ℃下200 r/min诱导24 h,收集发酵液测定酶活力和菌体生长量;在培养基中接种1%的种子液,培养不同时间(3、4、5、6、7 h)后,加入0.25 mmol/L的IPTG诱导剂,于15 ℃、200 r/min诱导24 h,收集发酵液测定酶活力和菌体生长量。
在上述最适条件下,以120 g/L果糖为底物进行全细胞催化30 min,整个体系经离心处理后0.22 µm滤膜过滤,滤液使用高效液相色谱(HPLC)检测,测定阿洛酮糖产量和酶活力,以未优化条件培养重组菌作为对照组。
2. 结果与分析
2.1 Cb-DAEase诱导表达及分离纯化
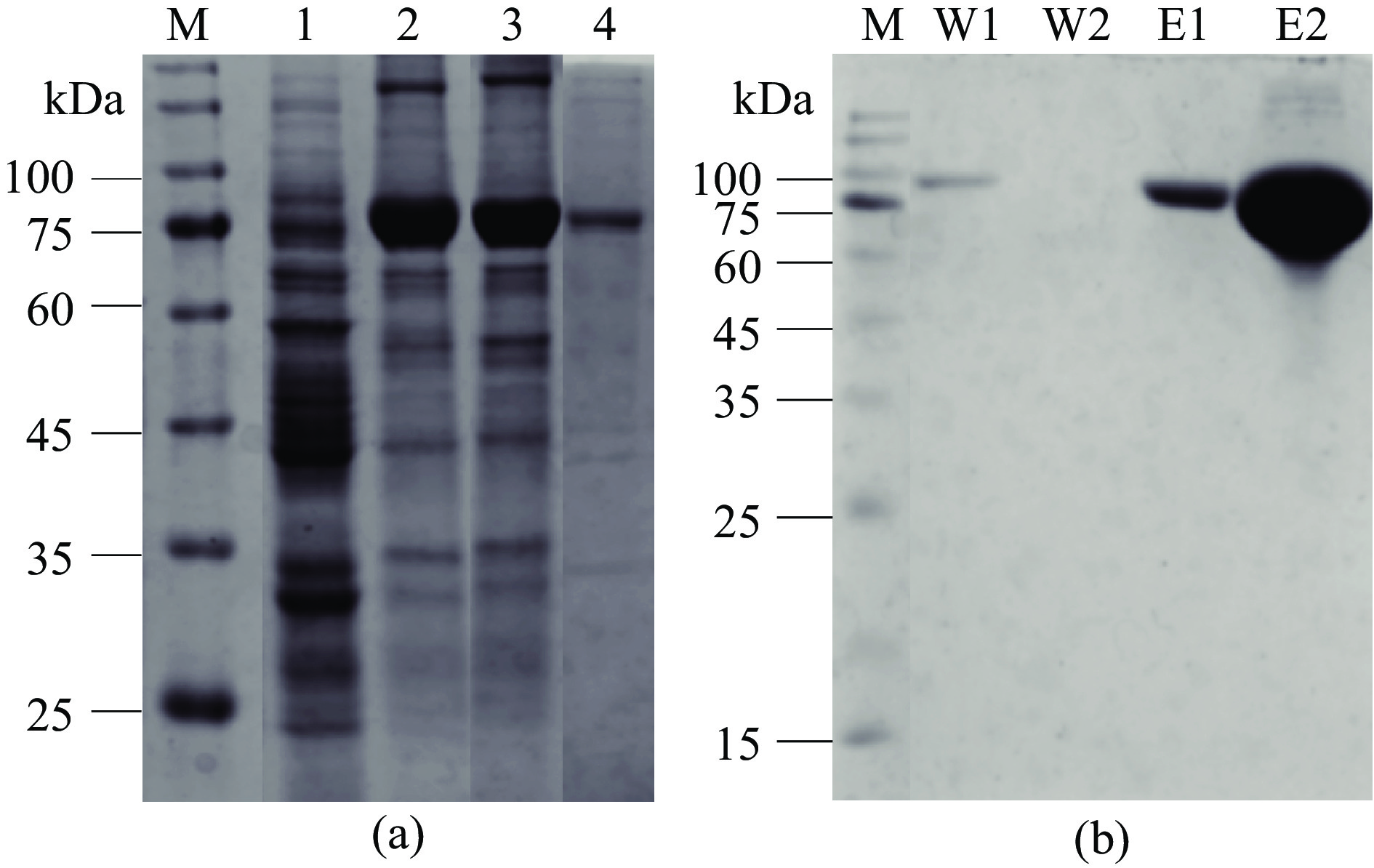
将构建成功的重组菌株BL21/TF-Cb接种于LB培养基中诱导表达Cb-DAEase,通过SDS-PAGE鉴定其分子量。DAEase的分子量大小约为34.2 kDa,携带TF分子伴侣的质粒分子量大小约为54 kDa,Cb-DAEase的分子量预测为88.2 kDa。如图1(a),在75~100 kDa之间有明显的条带,与预测值一致,说明重组质粒pCold TF-Cb-DAE成功在大肠杆菌宿主E. coli BL21(DE3)中表达。与总粗酶液(lane 2)相比,Cb-DAEase更多的以可溶性蛋白的方式存在于上清液中(lane 3),经分析软件Image J分析得出,上清中目的蛋白表达量占总蛋白的84.7%±1.4%,而沉淀中的包涵体含量仅占少部分(lane 4),说明冷休克表达质粒pCold TF在低温诱导条件下成功实现了DAEase在大肠杆菌中的高效可溶性表达。pCold载体上携带的分子伴侣TF可以伴随着蛋白质的初始折叠,通过与错误折叠的多肽暴露的疏水区域结合,并将分子转移到特定的亚细胞,从而避免蛋白质聚集,减少包涵体形成[21]。
![]() 图 1 大肠杆菌E. coli BL21(DE3)中表达Cb-DAEase的SDS-PAGE电泳图(a)以及纯化Cb-DAEase的SDS-PAGE结果(b)注:M:RealBand蛋白预染Marker;(a)泳道 1:阴性对照(BL21/TF);泳道 2:BL21/TF-Cb总粗酶液;泳道 3:BL21/TF-Cb上清(可溶);泳道 4:BL21/TF-Cb沉淀(包涵体);(b)泳道 W1:PBS+20 mmol/L咪唑洗脱液;泳道 W2:PBS+50 mmol/L咪唑洗脱液;泳道 E1、E2:纯化的Cb-DAEase。Figure 1. Expression of Cb-DAEase in E. coli BL21 (DE3) SDS-PAGE electrophoretogram (a) and SDS-PAGE results of purified Cb-DAEase (b)
图 1 大肠杆菌E. coli BL21(DE3)中表达Cb-DAEase的SDS-PAGE电泳图(a)以及纯化Cb-DAEase的SDS-PAGE结果(b)注:M:RealBand蛋白预染Marker;(a)泳道 1:阴性对照(BL21/TF);泳道 2:BL21/TF-Cb总粗酶液;泳道 3:BL21/TF-Cb上清(可溶);泳道 4:BL21/TF-Cb沉淀(包涵体);(b)泳道 W1:PBS+20 mmol/L咪唑洗脱液;泳道 W2:PBS+50 mmol/L咪唑洗脱液;泳道 E1、E2:纯化的Cb-DAEase。Figure 1. Expression of Cb-DAEase in E. coli BL21 (DE3) SDS-PAGE electrophoretogram (a) and SDS-PAGE results of purified Cb-DAEase (b)将重组菌株BL21/TF-Cb发酵上清液通过Ni Sepharose 6 Fast Flow亲和层析柱分离纯化目的蛋白,如图1(b)所示,条带W1为20 mmol/L洗脱液洗出的杂蛋白,条带W2为50 mmol/L洗脱液洗出的剩余杂蛋白(SDS-PAGE电泳图中未出现条带),表明20~50 mmol/L咪唑洗脱液条件下可将所有的杂蛋白洗脱。条带E1、E2为500 mmol/L洗脱液洗出的目的蛋白,纯化后的Cb-DAEase条带单一清晰占比高,回收的目的蛋白纯度高,可用于后续的酶学性质研究。
2.2 Cb-DAEase的酶学特性分析
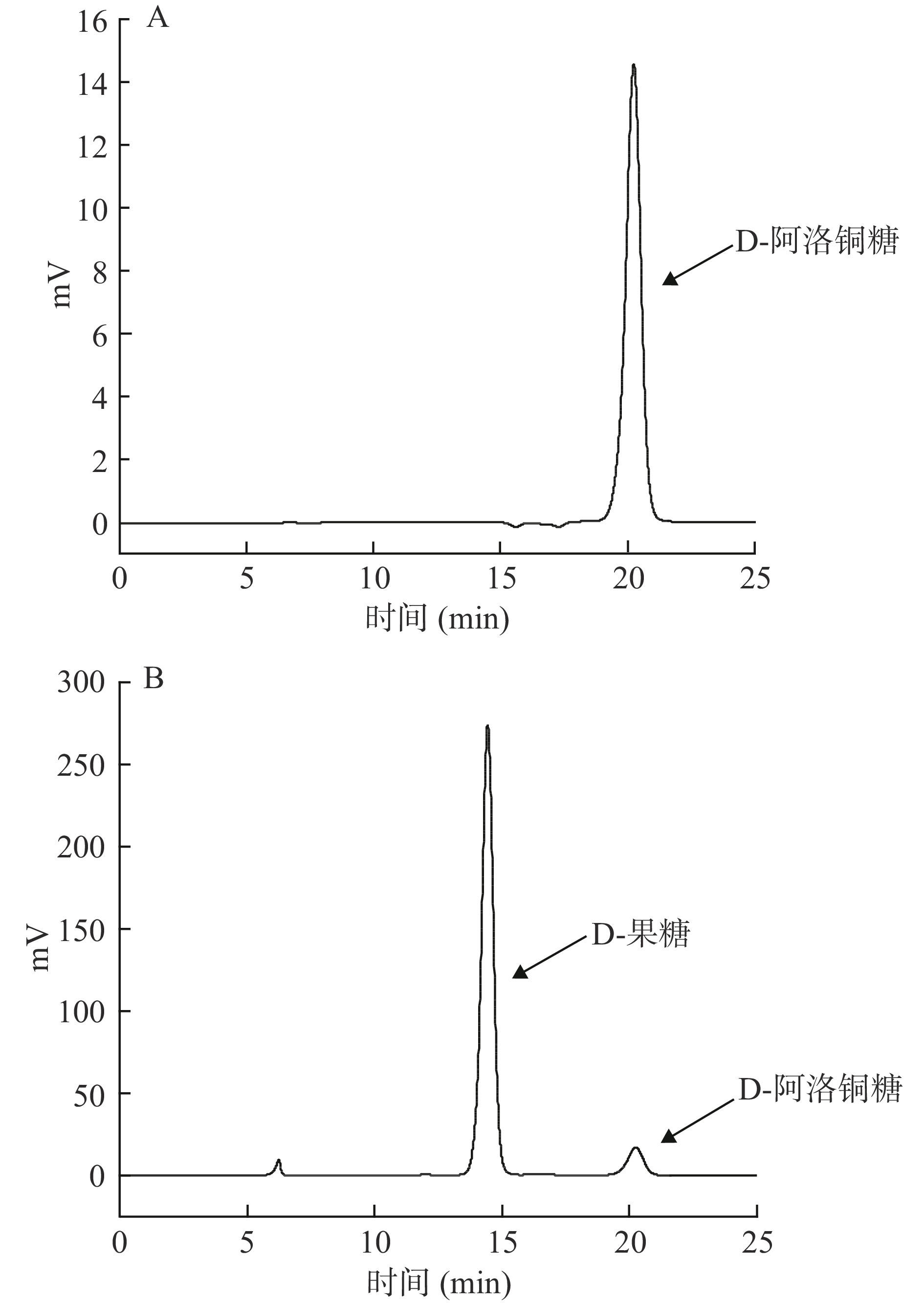
重组菌株在以D-果糖为底物的培养基中发酵,通过HPLC检测D-阿洛酮糖的生成。如图2所示,在20.25 min左右检测到D-阿洛酮糖,这表明Cb-DAEase可以在大肠杆菌冷休克系统中表达为活性蛋白,具有催化合成D-阿洛酮糖的能力。
![]() 图 2 重组Cb-DAEase活性HPLC检测注:A.D-allulose标准品;B.反应平衡后的产物。Figure 2. HPLC analysis of the bioactivity in recombinant Cb-DAEase
图 2 重组Cb-DAEase活性HPLC检测注:A.D-allulose标准品;B.反应平衡后的产物。Figure 2. HPLC analysis of the bioactivity in recombinant Cb-DAEase2.2.1 温度、pH、金属离子对Cb-DAEase酶活的影响
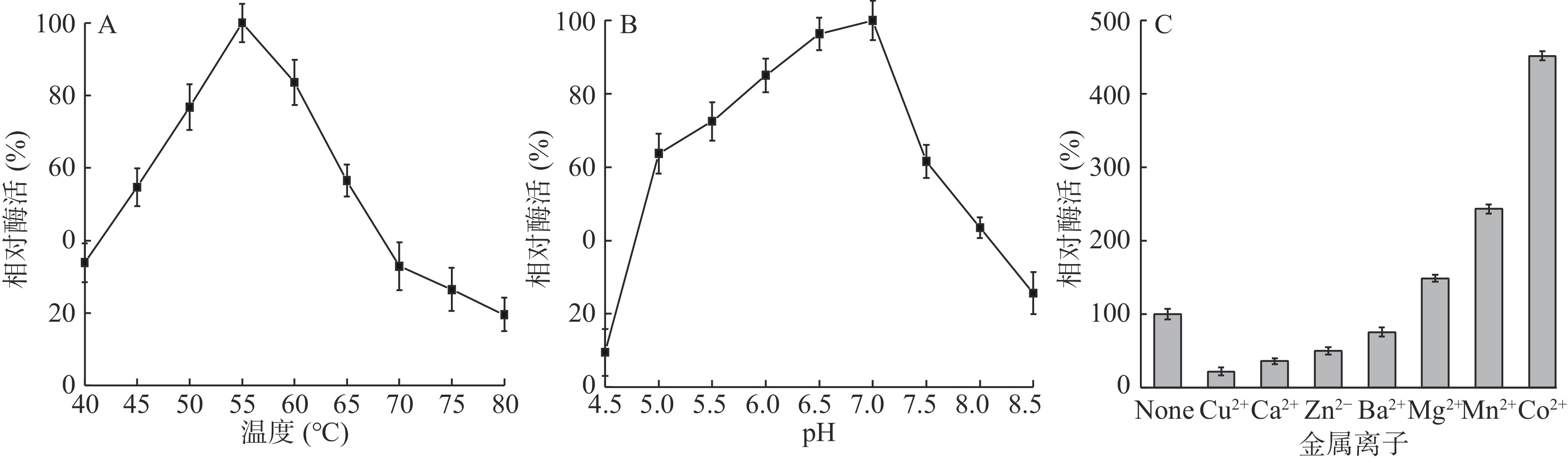
最适温度作为生物催化反应的重要参数,可反应其是否适用于工业化生产[23]。Cb-DAEase的最适反应温度为55 ℃(图3A)。在温度45~65 ℃内,Cb-DAEase仍保存50%以上的活性;但当温度超过55 ℃时,酶活力随着温度的升高而逐步下降,80 ℃时Cb-DAEase仅有19.56%的活性,说明温度是影响酶活力的主要因素。
![]() 图 3 重组Cb-DAEase的酶学性质注:A.重组酶最适反应温度的测定;B.重组酶的最适反应pH的测定;C.金属离子对重组酶活性影响的测定。Figure 3. Enzymatic properties of the recombinant Cb-DAEase
图 3 重组Cb-DAEase的酶学性质注:A.重组酶最适反应温度的测定;B.重组酶的最适反应pH的测定;C.金属离子对重组酶活性影响的测定。Figure 3. Enzymatic properties of the recombinant Cb-DAEaseCb-DAEase在pH7.0时酶活力最高(图3B),在pH5~7.5时Cb-DAEase可以保持60%以上的相对酶活力。Sato等[22]报道的Trpr-DAEase在pH 8.0时表现出最高活性,Zhu等[1]报道的重组DAEase在pH 7.5~8.0时活性最高,与目前发现的偏碱性DAEase不同,本研究的Cb-DAEase为中性酶,中性条件可避免褐变反应及其它非特异性副反应发生,减少后续分离纯化的难度,在工业应用中具有明显优势。
金属离子能够与酶形成络合物,稳定或破坏蛋白质的空间结构,在生物催化中起重要作用[23]。与对照组相比(图3C),金属离子Co2+、Mn2+及Mg2+在不同程度上提高了Cb-DAEase的活性,Mg2+将酶活力提高了1.48倍,Mn2+将酶活力提高了2.43倍,Co2+的激活作用最强,将酶活力提高了4.52倍,其激活作用原因可能是该阳离子在调节酶活力构象中起重要作用,从而促进果糖异构化成阿洛酮糖[24]。
2.3 碳源对细胞生长和Cb-DAEase活力的影响
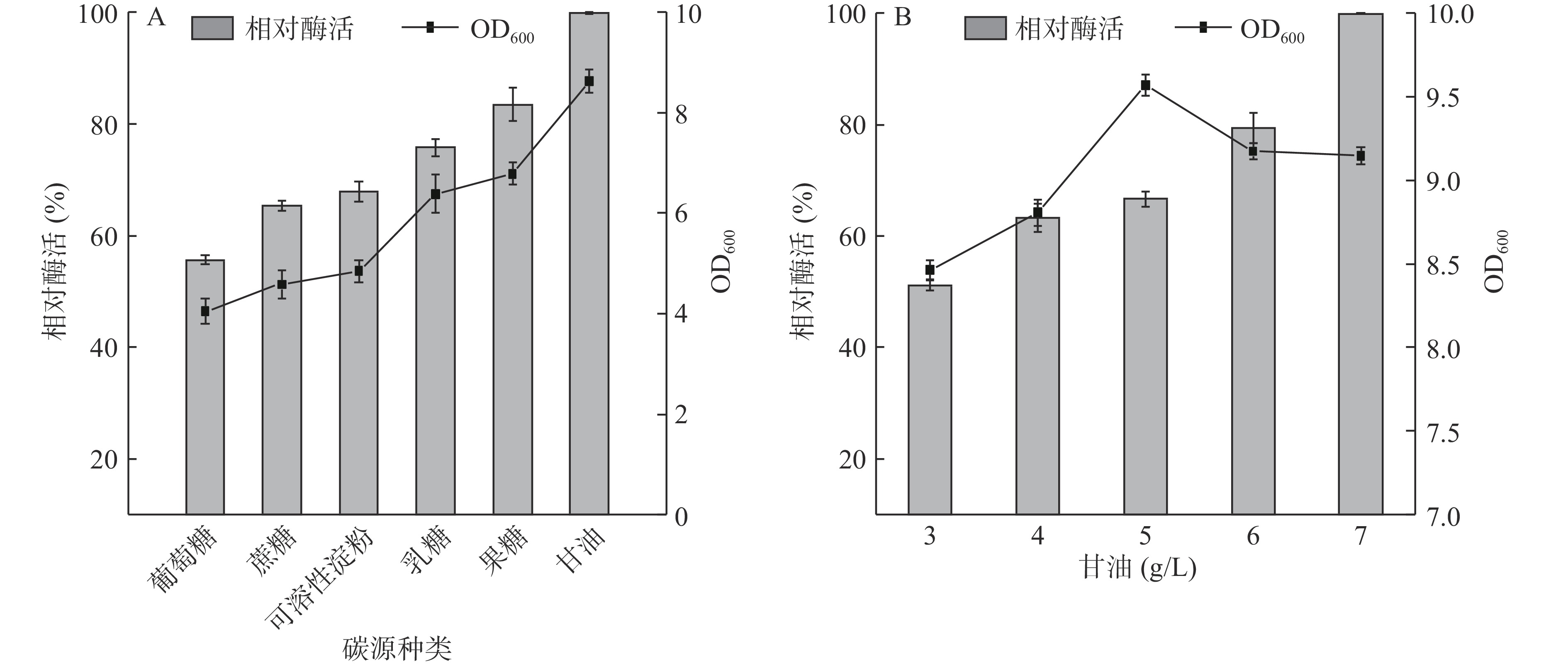
碳源是细胞生长的基础,其种类及浓度是影响胞内酶表达的重要因素[25]。如图4A,在添加葡萄糖、果糖、蔗糖、乳糖、可溶性淀粉和甘油的培养基中,以甘油为碳源时最有利于细胞生长、相对酶活最高,与Su[25]和Lei[26]等研究相似。甘油作为成本低廉的小分子稳定剂,可以降低发酵体系的极性,与酶蛋白形成氢键,有利于隔绝水分子环境及稳定蛋白结构[25]。
![]() 图 4 碳源种类及甘油质量浓度对菌体生长量和Cb-DAEase酶活性的影响Figure 4. Effects of carbon source type and glycerol concentration on growth mass and activity of Cb-DAEase
图 4 碳源种类及甘油质量浓度对菌体生长量和Cb-DAEase酶活性的影响Figure 4. Effects of carbon source type and glycerol concentration on growth mass and activity of Cb-DAEase通过在培养基中加入浓度为3~7 g/L的甘油,研究了不同甘油质量浓度对细胞生长和Cb-DAEase活性的影响。如图4B,当甘油浓度为3~5 g/L时,菌株生长量随着浓度的增加而提高;当甘油浓度为5~7 g/L时,菌株生长量随着浓度的增加而降低,这可能是由于碳代谢阻遏(CCR)效应[27],据报道,CCR效应会随着碳源浓度的增加而降低发酵培养基的pH,而且重组蛋白的表达通常会对细胞生长产生一定的代谢负担,降低细胞生物量[28]。Cb-DAEase的相对酶活力随着甘油浓度的增加而显著提高,7 g/L时相对酶活最高,菌体生长旺盛,故选择7 g/L甘油作为碳源。
2.4 氮源对细胞生长和Cb-DAEase活力的影响
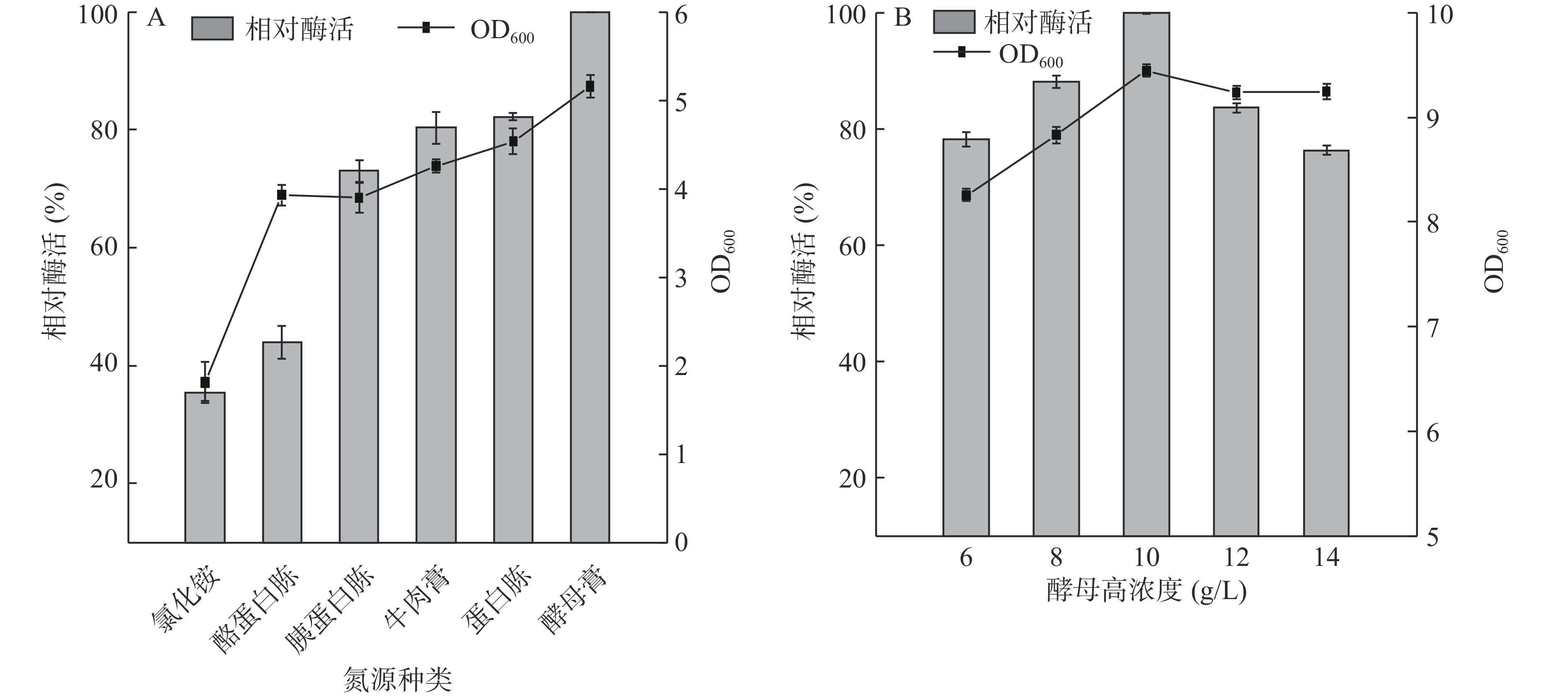
此外,本研究还考察了氮源种类对细胞生长和Cb-DAEase发酵活性的影响。在含有酵母膏、牛肉膏、蛋白胨和胰蛋白胨的培养基中,重组菌株生长量高,Cb-DAEase相对酶活在70%以上(图5A),其中酵母膏作为氮源时菌株生长量和相对酶活最高,说明有机氮源是基因工程菌表达Cb-DAEase的适宜氮源,这可能是由于有机氮源中除了含有蛋白质、肽及游离的氨基酸以外,还含有少量的维生素和生长因子,促进大肠杆菌的生长和蛋白表达[26,29]。
![]() 图 5 氮源种类及酵母膏质量浓度对菌体生长量和Cb-DAEase酶活性的影响Figure 5. Effects of nitrogen source type and yeast extract concentration on growth mass and activity of Cb-DAEase
图 5 氮源种类及酵母膏质量浓度对菌体生长量和Cb-DAEase酶活性的影响Figure 5. Effects of nitrogen source type and yeast extract concentration on growth mass and activity of Cb-DAEase酵母膏浓度在6~10 g/L的范围时,Cb-DAEase活力和细胞生长都随着浓度的增加而提高,10 g/L时酶活力和生物量最高(图5B),当酵母膏浓度在10~14 g/L的范围时,酶活力和细胞生长都随着浓度的增加而下降,故确定酵母膏的最适添加浓度为10 g/L。
2.5 培养条件对细胞生长和Cb-DAEase活力的影响
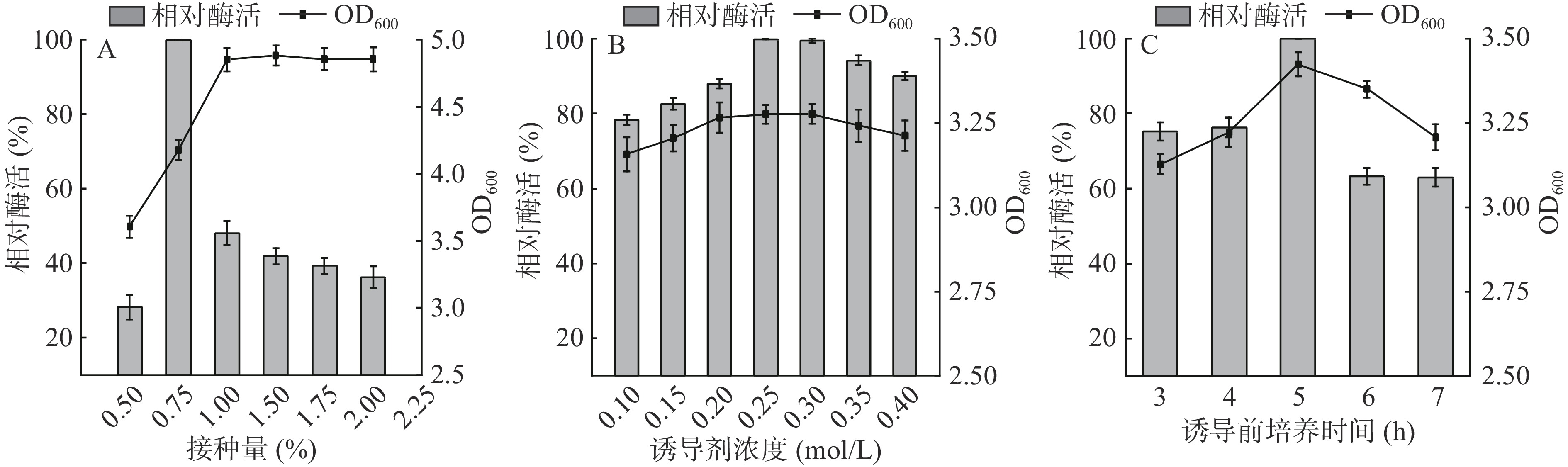
接种量是影响菌株生长快慢的因素之一,接种量过少会导致一个长时间的滞后期[30];增加接种量可促进微生物生长,缩短细胞生长的滞后期并降低感染杂菌的概率,但接种量过高会导致代谢废物增加,增加菌株的生物量,过早引起菌株之间的竞争[31],从而降低Cb-DAEase生物合成的速率,故研究接种量对细胞生长和Cb-DAEase活性的影响尤为重要。由图6A可知,Cb-DAEase的相对酶活随着接种量变化呈现先增加后降低的趋势,当接种量为1%时Cb-DAEase的相对酶活最高。
![]() 图 6 接种量(A)、诱导剂浓度(B)和诱导前培养时间(C)对生物量和Cb-DAEase表达的影响Figure 6. Effects of inoculum size (A), IPTG concentration (B) and induction time (C) on growth mass and expression of Cb-DAEase
图 6 接种量(A)、诱导剂浓度(B)和诱导前培养时间(C)对生物量和Cb-DAEase表达的影响Figure 6. Effects of inoculum size (A), IPTG concentration (B) and induction time (C) on growth mass and expression of Cb-DAEase以1%接种量为基础,改变诱导剂浓度,如图6B,随着诱导剂浓度的增加,酶活性和菌体生长量先增加后降低,在0.25 mmol/L时达到最高。IPTG浓度过高具有一定的毒性,导致菌体生长缓慢及形成包涵体,细胞内不溶性DAEase的比例增加[26];用量过多也会增加生产成本,故选择0.25 mmol/L作为最佳诱导剂浓度。
以上述条件为基础,改变诱导前培养时间,如图6C,培养时间在3~5 h时,酶活性和生长量随着培养时间的增加而提高;培养时间为5 h时,酶活力与生长量达到最大;此后延长培养时间,酶活力和生长量反而呈现下降趋势,说明培养5 h重组菌株生长旺盛,有利于产酶。因此,最终确定诱导前培养时间为5 h。
以120 g/L果糖作为底物,在上述条件下对重组菌株BL21/TF-Cb进行发酵优化培养,经全细胞反应催化后通过HPLC仪检测产物。Cb-DAEase活力达到(10.11±0.02) U/g,同比优化前(1.38±0.01) U/g提高了86.35%;阿洛酮糖产量为(11.47±0.04) g/L,同比优化前(1.03±0.02) g/L提高了91.02%,是Zhang等[31]发酵产量(4.56±0.01)g/L的2.51倍,优化效果十分显著,达到高效可溶性表达的目的,为工业化高产D-阿洛酮糖提供了数据支持。
3. 结论
在大肠杆菌中构建冷休克启动子诱导的外源蛋白表达系统,其新颖之处在于解决了胞内蛋白浓度过高导致折叠时间、空间受限而形成大量包涵体的问题,从而显著提高了DAEase在大肠杆菌中的表达水平。此外,以7 g/L甘油、10 g/L酵母膏、1%接种量、0.25 mmol/L IPTG、诱导前培养5 h的条件进行发酵显著降低了DAEase的生产成本, DAEase活力达到(10.11±0.02) U/g;以pH7.0、55 ℃和1 mmol/L Co2+催化条件进行全细胞反应,D-阿洛酮糖产量最高可达(11.47±0.04) g/L,基于这些结果,大肠杆菌冷休克表达系统在DAEase生产中显示出巨大的应用潜力,为进一步研究利用生物转化法生产D-阿洛酮糖提供技术参考。
-
![]()
图 1 大肠杆菌E. coli BL21(DE3)中表达Cb-DAEase的SDS-PAGE电泳图(a)以及纯化Cb-DAEase的SDS-PAGE结果(b)
注:M:RealBand蛋白预染Marker;(a)泳道 1:阴性对照(BL21/TF);泳道 2:BL21/TF-Cb总粗酶液;泳道 3:BL21/TF-Cb上清(可溶);泳道 4:BL21/TF-Cb沉淀(包涵体);(b)泳道 W1:PBS+20 mmol/L咪唑洗脱液;泳道 W2:PBS+50 mmol/L咪唑洗脱液;泳道 E1、E2:纯化的Cb-DAEase。
Figure 1. Expression of Cb-DAEase in E. coli BL21 (DE3) SDS-PAGE electrophoretogram (a) and SDS-PAGE results of purified Cb-DAEase (b)
![]()
图 2 重组Cb-DAEase活性HPLC检测
注:A.D-allulose标准品;B.反应平衡后的产物。
Figure 2. HPLC analysis of the bioactivity in recombinant Cb-DAEase
![]()
图 3 重组Cb-DAEase的酶学性质
注:A.重组酶最适反应温度的测定;B.重组酶的最适反应pH的测定;C.金属离子对重组酶活性影响的测定。
Figure 3. Enzymatic properties of the recombinant Cb-DAEase
![]()
图 4 碳源种类及甘油质量浓度对菌体生长量和Cb-DAEase酶活性的影响
Figure 4. Effects of carbon source type and glycerol concentration on growth mass and activity of Cb-DAEase
![]()
图 5 氮源种类及酵母膏质量浓度对菌体生长量和Cb-DAEase酶活性的影响
Figure 5. Effects of nitrogen source type and yeast extract concentration on growth mass and activity of Cb-DAEase
-
[1] ZHU Yueming, MEN Yan, BAI Wei, et al. Overexpression of D-psicose 3-epimerase from Ruminococcus sp. in Escherichia coli and its potential application in D-psicose production[J]. Biotechnology Letters,2012,34(10):1901−1906. doi: 10.1007/s10529-012-0986-4
[2] WEI Hongbei, ZHANG Ruoxuan, WANG Leyi, et al. Expression of D-psicose-3-epimerase from Clostridium bolteae and Dorea sp. and whole-cell production of D-psicose in Bacillus subtilis[J]. Annals of Microbiology,2020,70(1):403−410.
[3] ZHANG Wenli, LI Hao, JIANG Bo, et al. Production of D-allulose from D-glucose by Escherichia coli transformant cells co-expressing D-glucose isomerase and D-psicose 3-epimerase genes[J]. Journal of the Science of Food and Agriculture,2017,97(10):3420−3426. doi: 10.1002/jsfa.8193
[4] JUNEJA A, ZHANG G, JIN Y S, et al. Bioprocessing and technoeconomic feasibility analysis of simultaneous production of D-psicose and ethanol using engineered yeast strain KAM-2GD[J]. Bioresource Technology, 2019, 275: 27−34.
[5] HU Mengying, LI Mengli, JIANG Bo, et al. Bioproduction of D-allulose: Properties, applications, purification, and future perspectives[J]. Comprehensive Reviews in Food Science and Food Safety,2021,20(6):6012−6026. doi: 10.1111/1541-4337.12859
[6] LI Can, GAO Ling, DU Kai, et al. Production of D-allulose from D-fructose using immobilized L-rhamnose isomerase and D-psicose 3-epimerase[J]. Bioprocess and Biosystems Engineering,2020,43(4):645−653. doi: 10.1007/s00449-019-02262-y
[7] ZHANG Wenli, LI Hao, ZHANG Tao, et al. Characterization of a D-psicose 3-epimerase from Dorea sp. CAG317 with an acidic pH optimum and a high specific activity[J]. Journal of Molecular Catalysis B: Enzymatic,2015,120:68−74. doi: 10.1016/j.molcatb.2015.05.018
[8] PARK C S, PARK C S, Shin K C, et al. Production of D-psicose from D-fructose by whole recombinant cells with high-level expression of D-psicose 3-epimerase from Agrobacterium tumefaciens[J]. Journal of Bioscience and Bioengineering,2016,121(2):186−190. doi: 10.1016/j.jbiosc.2015.06.010
[9] 苏鹏, 龚国利. 优化大肠杆菌表达外源蛋白的研究进展[J]. 生物技术通报,2017,33(2):16−23. [SU P, GONG G L. Research progress on optimizing the expression of exogenous proteins in Escherichia coli[J]. Biotechnology Bulletin,2017,33(2):16−23. doi: 10.13560/j.cnki.biotech.bull.1985.2017.02.003 [10] 马媛媛, 何健民, 康永杰. 外源性蛋白在大肠杆菌中可溶性表达的策略综述[J]. 世界科技研究与发展,2015,37(5):627−630. [MA Y Y, HE J M, KANG Y J. Reviews on strategies of enhance heterologous protein soluble expression in Escherichia coli[J]. World Sci-Tech R & D,2015,37(5):627−630. doi: 10.16507/j.issn.1006-6055.2015.05.033 [11] DOMINIC E, DEB K C. Enhancement of soluble protein expression through the use of fusion tags[J]. Current Opinion in Biotechnology,2006,17(4):353−358. doi: 10.1016/j.copbio.2006.06.003
[12] TSENG W C, HSU C T, CHANG H C, et al. Fusion of the peptide derived from the acidic tail of alpha-synuclein improves the thermostability and soluble expression of recombinant Agrobacterium sp. D-allulose 3-epimerase[J]. Biochemical Engineering Journal,2021,165:107828. doi: 10.1016/j.bej.2020.107828
[13] CHESSHYRE J A, HIPKISS A R, et al. Low temperatures stabilize interferon α-2 against proteolysis in Methylophilus methylotrophus and Escherichia coli[J]. Applied Microbiology & Biotechnology,1989,31(2):158−162.
[14] KAE H C, TAKASHI Y B, MASAHIRO I, et al. Cold shock proteins improve E. coli cell-free synthesis in terms of soluble yields of aggregation-prone proteins[J]. Biotechnology and Bioengineering,2020,117(6):1628−1639. doi: 10.1002/bit.27326
[15] 闫真, 聂尧, 徐岩. 重组大肠杆菌表达氧化还原酶不对称还原2-羟基苯乙酮的研究[J]. 工业微生物,2011,41(5):1−5. [YAN Z, NIE R, XU Y. Asymmetric reduction of 2-hydroxyacetophenone by recombinant Escherichia coli expressing oxidoreductase[J]. Industrial Microbiology,2011,41(5):1−5. doi: 10.3969/j.issn.1001-6678.2011.05.001 [16] NENG Xiong, DONG Xie, YAN Dong, et al. Efficient biosynthesis of 1-cyanocyclohexaneacetic acid using a highly soluble nitrilase by N-terminus modification of novel peptide tags[J]. Biochemical Engineering Journal,2021,176:108207. doi: 10.1016/j.bej.2021.108207
[17] QING Guoliang, MA Lichung, KHORCHID A, et al. Cold-shock induced high-yield protein production in Escherichia coli[J]. Nature Biotechnology,2004,22(7):877−882. doi: 10.1038/nbt984
[18] ZHAO Jingyi, CHEN Jing, WANG Huiyi, et al. Enhanced thermostability of D-psicose 3-epimerase from Clostridium bolteae through rational design and engineering of new disulfide bridges[J]. International Journal of Molecular Sciences,2021,22(7):10007.
[19] MIN J S, EUN R K, SANG J K, et al. D-allulose production from D-fructose by putative dolichol phosphate mannose synthase from Bacillus sp. with potential D-allulose 3-epimrase activity[J]. Biotechnology and Bioprocess Engineering,2021,26(6):976−984. doi: 10.1007/s12257-021-0007-3
[20] YANG Lin, CHEN Tianjiao, WANG Fen, et al. Structures of β-glycosidase LXYL-P1-2 reveals the product binding state of GH3 family and a specific pocket for taxol recognition[J]. Communications Biology,2020,3(1):1−8. doi: 10.1038/s42003-019-0734-6
[21] 朱星星, 杨培周, 杜明睿, 等. 根癌农杆菌D-阿洛酮糖-3-差向异构酶基因克隆、结构预测及原核表达[J]. 食品科学技术学报,2019,37(3):61−66. [ZHU X X, YANG P Z, DU M R, et al. Gene cloning, structural prediction, and prokaryotic expression of Agrobacterium tumefaciens D-psicose-3-epimerase[J]. Journal of Food Science and Technology,2019,37(3):61−66. doi: 10.3969/j.issn.2095-6002.2019.03.008 [22] SATO K, KANEKO T, MIURA T Y, et al. Purification and characterization of a milk-clotting enzyme from Hericium erinaceum[J]. Food Science and Technology Research,2019,25(5):735−741. doi: 10.3136/fstr.25.735
[23] MAHBOOBEH N, AMIR-HASSAN Z, ROYA G, et al. Optimized protocol for soluble prokaryotic expression, purification and structural analysis of human placenta specific-1(PLAC1)[J]. Protein Expression and Purification,2017,133:139−151. doi: 10.1016/j.pep.2017.03.011
[24] CHEN Xiaoyan, WANG Wen, XU Jingliang, et al. Biochemical characterization of a D-psicose 3-epimerase from Treponema primitia ZAS-1 and its application on enzymatic production of d-psicose[J]. Journal of the Science of Food and Agriculture,2016,96(1):49−56. doi: 10.1002/jsfa.7187
[25] SANTA-BELL E, KOVACS N K, ALACS B, et al. Immobilization of phenylalanine ammonia-lyase via EDTA based metal chelate complexes-optimization and prospects[J]. Periodica Polytechnica-Chemical EngineerinG,2021,65(3):308−319. doi: 10.3311/PPch.17891
[26] GU Lei, ZHANG Juan, LIU Baihong, et al. High-level extracellular production of D-psicose-3-epimerase with recombinant Escherichia coli by a two-stage glycerol feeding approach[J]. Bioprocess and Biosystems Engineering,2013,36(11):1767−1777. doi: 10.1007/s00449-013-0952-0
[27] 孙帆, 宿玲恰, 张康, 等. D-阿洛酮糖3-差向异构酶在枯草芽孢杆菌中的高效表达及固定化细胞研究[J]. 中国生物工程杂志,2018,38(7):83−88. [SUN F, SU L Q, ZHANG K, et al. D-psicose 3-epimerase gene overexpression in Bacillus subtilis and immobilization of cells[J]. China Biotechnology,2018,38(7):83−88. doi: 10.13523/j.cb.20180711 [28] SU Lingqia, SUN Fan, LIU Zhanzhi, et al. Highly efficient production of Clostridium cellulolyticum H10 D-psicose 3-epimerase in Bacillus subtilis and use of these cells to produce d-psicose[J]. Microbial Cell Factories,2018,17(1):188. doi: 10.1186/s12934-018-1037-1
[29] 孙丽慧, 王云晓, 吕诗文, 等. 1株高产L-乳酸菌株的分离鉴定及其发酵培养基优化[J]. 食品科学,2018,39(6):135−140. [SUN L H, WANG Y X, LÜ S W, et al. Isolation and identification of a strain for high-yield production of L-lactic acid and optimization of its culture medium[J]. Food Science,2018,39(6):135−140. doi: 10.7506/spkx1002-6630-201806022 [30] GUO, Fenfen, LI Xuezhi, ZHAO Jian, et al. Optimizing culture conditions by statistical approach to enhance production of pectinase from Bacillus sp. Y1[J]. BioMed Research International,2019,19:1−10.
[31] ZHANG Jun, XU Chao, CHEN Xiaoyan, et al. Engineered Bacillus subtilis harbouring gene of D-tagatose 3-epimerase for the bioconversion of D-fructose into D-psicose through fermentation[J]. Enzyme and Microbial Technology,2020,136:109531. doi: 10.1016/j.enzmictec.2020.109531
 下载:
下载:





 下载:
下载:

计量
- 文章访问数:
- HTML全文浏览量:
- PDF下载量:
