


Research Progress of Ectopic Olfactory Trace Amine-associated Receptors
-
摘要: 痕量胺相关受体(trace amine-associated receptors, TAARs)是一类G蛋白偶联受体,除TAAR1外,其余TAARs与嗅觉受体一样都在嗅觉上皮表达并发挥感知气味的作用。近年来,非嗅觉组织中同样发现TAARs高表达,提示TAARs可能存在重要的异位生理功能。事实上,相关研究证实内源性以及外源性特定痕量胺通过作用于不同的TAARs在非嗅觉组织中参与调节多种生理功能,表明TAARs有成为新的诊断和治疗靶点的潜力。本文系统介绍了异位嗅觉TAARs的表达、内源性以及外源性的配体、食物中的生物胺、异位嗅觉TAARs介导的信号通路以及生理和病理功能的现有研究,一方面为未来药物靶点开发、生理病理学研究提供新思路,另一方面也为食品中生物胺的生物活性研究提供新的研究方向。
-
关键词:
- 异位嗅觉痕量胺相关受体 /
- 食物生物胺 /
- 非嗅觉组织 /
- G蛋白偶联受体
Abstract: Trace amine-associated receptors (TAARs) are a class of G-protein-coupled receptors, and with the exception of TAAR1, the remaining TAARs, like the olfactory receptors, are expressed in the olfactory epithelium and function to sense odorants. In recent years, the high expression of TAARs has been found in non-olfactory tissues, suggesting that TAARs may have important ectopic physiological functions. Recent studies have confirmed that endogenous and exogenous specific trace amines regulate a variety of physiological functions in non-olfactory tissues by acting on different TAARs, indicating that TAARs have the potential to become a new diagnostic and therapeutic target. This paper systematically introduces the expression of ectopic olfactory TAARs, endogenous and exogenous ligands, biogenic amines in food, ectopic olfactory TAARs-mediated signal pathways, and physiological and pathological functions. On the one hand, it provides new ideas for the future drug target development, physiological and pathological research, on the other hand, it also provides new research ideas for the biological activity of biogenic amines in food. -
G蛋白偶联受体(G Protein-Coupled Receptors,GPCRs)是一种七次跨膜受体,通过响应各种内源性和外源性神经递质、肠道微生物代谢产物、气味分子、生物胺等多种配体,偶联到不同的异源三聚体G蛋白,发挥重要的信号传导作用[1-2]。由于GPCRs在全身大多数组织中均有表达,能够调节机体生理现象的多个方面,所以是多种疾病的重要治疗靶点[3]。嗅觉受体(Olfactory Receptors,ORs)是一类特殊的GPCRs,主要表达在嗅觉感受神经元(Olfactory Sensory Neuron, OSN)中,可以被特定的气味分子激活,并通过鼻腔中的嗅觉系统产生的电信号传向大脑感知外界的化学信号[4]。近年来研究发现ORs不仅仅只在OSN中表达,在非嗅觉组织中也有众多的ORs存在,并发挥重要的生理功能[5-8]。
相较于ORs而言,TAARs被证实是进化保守的嗅觉受体小家族,除TAAR1外,其他的TAARs主要分布于嗅上皮OSN中,先前研究多是集中于嗅觉TAARs在感知气味分子的作用。在OSN中TAARs和ORs一样,TAARs也是单等位基因表达,即表达TAARs等位基因的感觉神经元不表达其他TAARs或OR,遵循“一个神经元,一个受体”的规则[9-10]。嗅觉TAARs还可触发与ORs相同的下游级联信号,表达经典的Golf/cAMP信号通路[9,11]。Borowsky等最先于2001年研究发现,因两个TAAR1/TAAR4受体识别内源性痕量胺,因此被定义为“痕量胺受体”[12-13]。近年来研究发现除嗅上皮OSN外,TAARs也广泛分布于多种外周组织,提示TAARs具有潜在的异位作用。事实上,有研究已经就TAARs在异位组织(免疫系统、内分泌系统、心血管系统)的多种生理作用进行了报道[14-16]。
本文将系统介绍异位嗅觉TAARs的表达、内源性以及外源性的配体、食物中的生物胺、异位嗅觉TAARs介导的信号通路以及生理和病理功能的现有研究,一方面为未来药物靶点开发、生理病理学研究提供新思路,另一方面也为食品中生物胺的生物活性研究提供新的研究思路。
1. 异位嗅觉TAARs及其配体
1.1 异位嗅觉TAARs的表达
TAARs是一类在脊椎动物中保守的GPCRs,Buck等[9]在2006年发现TAARs是一种新型的小家族嗅觉受体,属于嗅觉上皮中进化保守的化学感觉受体。在人类中有6个功能性TAAR基因,小鼠中有15个功能性TAAR基因,大鼠中有16个功能性TAAR基因[17-18]。除TAAR1外,所有TAARs都可以作为嗅觉受体,感知气味,特别是挥发性胺,因此被称为“嗅觉痕量胺相关受体”。尽管,最初人们认为嗅觉TAARs仅局限于嗅上皮OSN,然而,随着研究的深入,科学家们发现嗅觉TAARs广泛分布于全身组织[19-21],例如,TAAR2、TAAR6和TAAR9已在小鼠十二指肠粘膜细胞和各种人类白细胞群中鉴定发现[22-23]。TAAR5、TAAR6和TAAR8在不同的大脑区域表达,包括杏仁核、基底节、额叶皮质、海马和黑质等[12,20,24]。据报道,心脏、肾脏、肺、肌肉、脾脏、胃、脊髓和睾丸也都表达各种TAARs[12,21,25-27](表1)。
表 1 TAARs及其内源性配体Table 1. TAARs and their endogenous ligands受体类型 种属 主要表达部位 检测方法 配体 参考文献 TAAR1 人、大鼠、小鼠 脑、脊髓、胰腺β细胞、胃、肠道、白细胞 RT-PCR β-苯乙胺、
p-酪胺[23,42-44] TAAR2 人、大鼠、小鼠 肠道、心脏、睾丸、白细胞 RT-PCR / [23,45-47] TAAR3 大鼠、小鼠 下丘脑、胰腺β细胞 RT-PCR 异戊胺 [48] TAAR4 大鼠、小鼠 大脑、胰腺β细胞 RT-PCR β-苯乙胺 [14] TAAR5 人、大鼠、小鼠 大脑、脊髓、肠道、睾丸、白细胞 RT-PCR 三甲胺 [49−50] TAAR6 人、大鼠、小鼠 大脑、肠道、睾丸、白细胞、肾脏 RT-PCR / [19,47,51] TAAR7a 大鼠、小鼠 大脑、肾脏 RT-PCR、
微阵列/ [52] TAAR7b 大鼠 大脑、胰腺β细胞 N,N-二甲基烷基胺 [14,52] TAAR7c 大鼠、小鼠 大脑、胰腺β细胞 / TAAR7d 大鼠、小鼠 大脑、胰腺β细胞 N,N-二甲基烷基胺 TAAR7e 大鼠、小鼠 大脑、胰腺β细胞 N,N-二甲基烷基胺 TAAR7f 小鼠 大脑 N,N-二甲基烷基胺 TAAR7g 大鼠 大脑、胰腺β细胞 / TAAR7h 大鼠 大脑、胰腺β细胞 N,N-二甲基烷基胺 TAAR8a 大鼠、小鼠 星形胶质细胞、嗅觉上皮、杏仁核、白细胞、
胃、肠、心脏、睾丸、肺、脾、肾、肌肉RT-PCR N-甲基哌啶 [46,53] TAAR8b 大鼠、小鼠 TAAR8c 大鼠、小鼠 TAAR9 人、大鼠、小鼠 嗅上皮、脊髓、肠、脾、骨骼肌、垂体、白细胞 RT-PCR N-甲基哌啶 [19,42] 1.2 机体中内源性生物胺
机体内的经典的生物胺包括组胺、多巴胺以及肾上腺素等,主要是由神经元中氨基酸脱羧产生,通常作为化学信使在中枢或外周组织与细胞表面GPCRs结合发挥重要生理功能。机体内的痕量胺包括机体内芳香族氨基酸(即:苯丙氨酸、酪氨酸、色氨酸)代谢产物,包括酪胺、β-苯乙胺、色胺、章胺和3-碘甲状腺原氨酸等(表1)。章胺是一种羟基苯乙胺,主要存在于昆虫、软体动物和其他无脊椎动物中,首次是从章鱼的唾液腺中分离出来[28-29]。β-苯乙胺是哺乳动物大脑中苯丙氨酸脱羧产生的一种内源性胺,β-苯乙胺分布在哺乳动物的整个大脑,但在多巴胺能区尾壳核的浓度相对较高[30-32]。色胺是一种单胺类生物碱,含有吲哚环结构,结构类似于色氨酸,哺乳动物的大脑中发现了微量的色胺,发挥神经调节剂或神经递质的作用[33]。去氧肾上腺素是一种生物碱,自然存在于一些动物中,在正常的人类尿液和其他哺乳动物组织中也发现了低水平的脱氧肾上腺素[34-35]。3-碘甲状腺原氨酸是一种内源性甲状腺胺,通过质谱法、高效液相色谱法等检测技术,发现在大脑、心脏以及血液中都检测到了3-碘甲状腺原氨酸的存在[36-37]。3-甲氧基酪胺是由神经递质多巴胺在儿茶酚-O-甲基转移酶代谢下生成的代谢产物,在单胺氧化酶代谢作用下形成高香草酸,通常会随尿液排出体外[38]。N-甲基酪胺,又称4-羟基-N-甲基苯乙胺,是人类体内的一种微量胺神经调节剂,由微量胺苯乙胺衍生而来[39-40]。此外,在人的尿液中还检测到N-甲基苯乙胺,研究发现N-甲基苯乙胺是由苯乙胺作为底物,在苯乙醇胺N-甲基转移酶的代谢作用下产生[41]。三甲胺是机体内一类经过黄素单氧酶3与肠道菌群共同代谢的产物。但是,值得注意的是,除上述所描述的各种不同的化合物成为TAARs的配体外,还有多个TAARs为孤儿受体,现有研究暂未发现有天然配体,提示我们仍有大量的研究亟待开展。
1.3 食物中外源性生物胺
TAARs的配体来源除了机体内的痕量生物胺外,还有一些来自外源的食物或天然产物。例如,酪胺是一种由动物、植物和微生物中的氨基酸酪氨酸脱羧而产生的生物胺(表2),可能存在于几乎所有含有蛋白质或游离酪氨酸的食物中。Tarján等[54]研究显示蔬菜中甜菜根、卷心菜、甘蓝、花椰菜、黄瓜、萝卜中都含有丰富的酪胺。Andersen等[55]对奶制品中的酪胺含量进行检测发现芝士和牛奶中都含有大量的酪胺,其中芝士中相对含量较高。此外,在鱼类中也存在有大量的酪胺[56]。微生物代谢产物也是微量胺的重要来源,其中酪胺、苯乙胺等都是重要的细菌代谢产物。例如,Gale等[57]研究发现粪肠球菌可将酪氨酸代谢成为一种重要的TAARs配体——酪胺。此外,肉食杆菌属菌株是一类耐盐的嗜冷细菌,能够在恶劣的生长条件下生长,大多数肉杆菌属菌株都可以产生大量的酪胺和苯乙胺[58]。Moreno等[59]从葡萄酒中分离到几株产酪胺的乳杆菌。β-苯乙胺也存在于各种食品中,如巧克力[60-62]、奶酪[63]和葡萄酒[64-65],其中在巧克力中的含量相对较高[42]。
表 2 食物中的生物胺及其受体Table 2. Biogenic amines and their receptors in food生物胺 受体类型 主要食物来源 参考文献 酪胺 TAAR1 牛奶以及奶制品(酪乳、牛奶、酸奶) [29,42,55,57,59] 芝士 肉类(鸡肝、猪肉、火腿) 鱼(鳕鱼、沙丁鱼、马鲛鱼、金枪鱼、虾) 水果(牛油果、香蕉、葡萄、柑橘、西瓜、覆盆子) 蔬菜(甜菜根、卷心菜、胡萝卜、花椰菜、黄瓜、莴苣、
甜红椒、大豆、菠菜、西红柿、土豆)β-苯乙胺 TAAR1
TAAR4奶酪、葡萄酒、巧克力 [42] 三甲胺 TAAR5 猪肉、鱼、虾 [69−70] 腐胺 / 肉类、腌制鱼类、腌制蔬菜 [66,68, 72] 尸胺 TAAR13c 肉类、腌制鱼类、腌制蔬菜 [66,68,72,77] 组胺 TAAR1 肉类、腌制鱼类、腌制蔬菜 [66,68,72,78] 亚精胺 / 猪肉、羊肉、牛肉、鸡肉 [71] 精胺 TAAR348 猪肉、羊肉、牛肉、鸡肉 [71,79] 色胺 TAAR1 烤肉、干发酵香肠 [80] 肉制品中因为含有丰富的蛋白质,该类食物可以在细菌暴露的条件下进行发酵产生多种生物胺,例如,常见的组胺、亚精胺、精胺、色胺、苯乙胺、酪胺等都是在食物中可以检测到的含量较高的生物胺[66]。研究发现,肉制品中的腐胺、组胺、酪胺等含量较高,产生该种生物胺的微生物主要有假单胞杆菌属、乳酸杆菌属、肠杆菌属等。此外,在不同种类的肉食食品中,生物胺的含量也有一定的区别,例如,在猪肉、羊肉、牛肉、鸡肉等新鲜原料肉中精胺和亚精胺含量较高,其他生物胺类物质在新鲜原料肉中含量则相对较低[67]。不同的水产品因为含有蛋白质的种类、数量以及储藏条件等有一定的差异,所生成的特征生物胺的种类和数量也会有所不同,例如,常温下腌制的带鱼以及金线鱼中以组胺、腐胺、尸胺为主,去内脏、头尾之后的马鲛鱼以及秋刀鱼在-30 ℃储存条件下产生的生物胺以苯乙胺、尸胺、组胺为主[68]。三甲胺则主要存在于腐败的鱼虾以及肉制品中[69-70],鱼肉中的氧化三甲胺在微生物代谢酶的作用下产生三甲胺,并随着储存时间的延长,三甲胺的含量也逐渐升高,是鱼类等食物腐败的标志物[71]。发酵食品在发酵过程中通常会伴随微生物的迅速生长以及游离氨基酸的产生,所以会产生多种生物胺,例如,腐胺、尸胺、组胺以及酪胺是东北酸菜以及现代工业泡菜半成品中主要的生物胺,其中以尸胺的含量最为显著。此外,不同的蔬菜类型在发酵的过程中产生的生物胺的类型也有一定的差异,例如,腌制萝卜中的生物胺检出量较低,而泡豇豆中则含有较高水平的生物胺[72]。
食物中外源直接摄入的生物胺或者其前体物质在机体内可代谢生成生物胺,适量外源摄入的生物胺可与TAARs结合发挥相关的生理作用,随后在机体的代谢作用下排出体外,对身体健康有益。然而摄入过量的生物胺或者机体代谢能力不足时就会对机体造成损伤,引发中毒导致不良反应。例如,过量的组胺会引起头疼、心悸、呕吐以及血压异常等不良反应,此外还伴随有一定的神经毒性[73-74];摄入过多的酪胺、色胺或者β-苯乙胺则会引起头疼以及高血压等[55];腐胺和尸胺则具有强烈的刺激性,进入机体后可抑制组胺和酪胺相关代谢酶的活性,进一步增强组胺和酪胺的毒性作用,此外,腐胺和尸胺还可与亚硝酸盐反应生成强致癌性的亚硝胺,增加诱发癌症的风险[75-76]。
2. 异位嗅觉TAARs信号传导通路
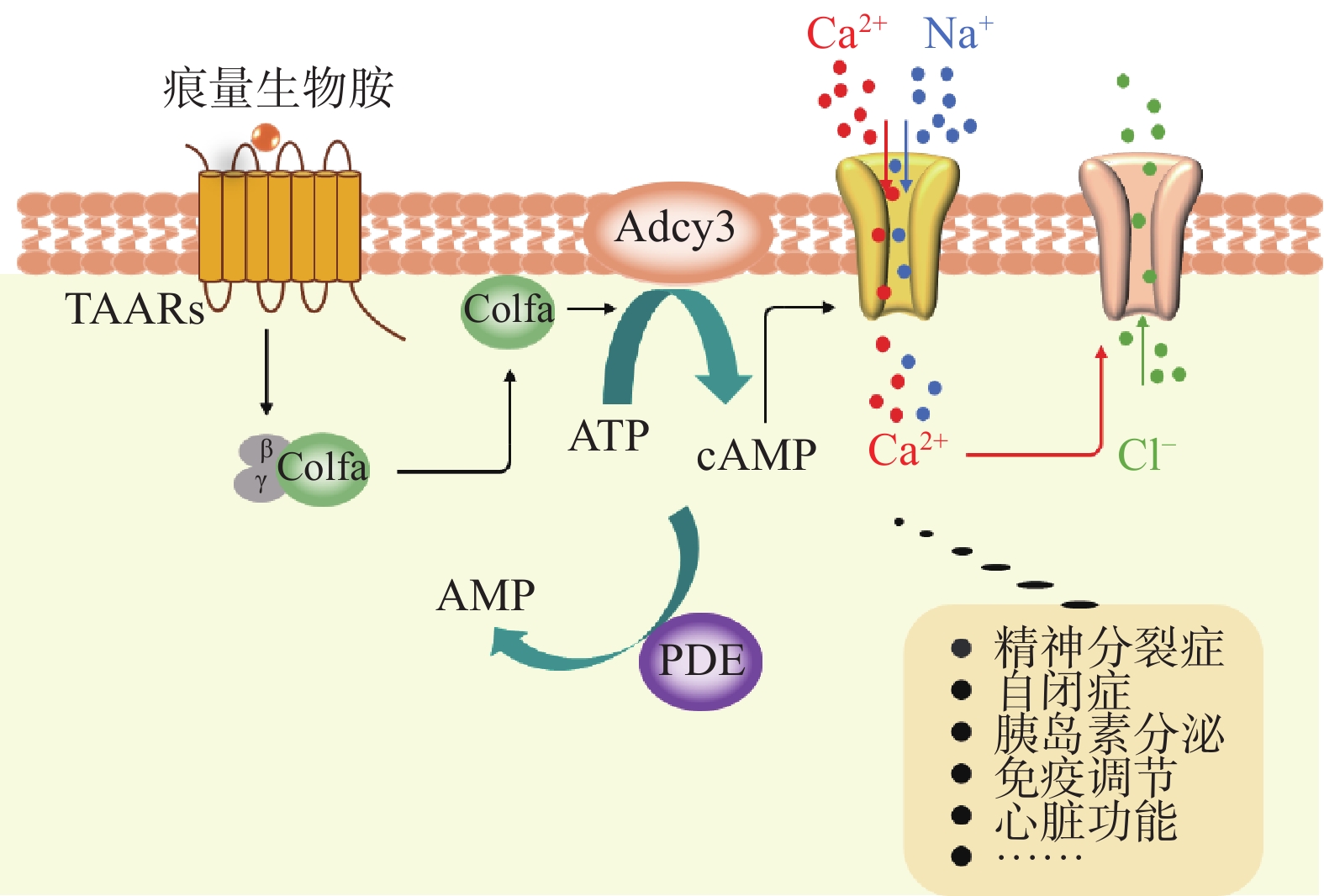
嗅觉TAARs通过典型的嗅觉信号转导级联来转导它们各自的配体,嗅觉受体结合嗅觉特异性G蛋白(Golf),激活腺苷酸环化酶(Adenylate cyclase 3, Adcy3),在Adcy3的作用下腺嘌呤核苷三磷酸(Adenosine triphosphate, ATP)转化为细胞内重要的第二信使-环磷酸腺苷(Cyclic adenosine 3’, 5’-monophosphate, cAMP),cAMP浓度升高后会使细胞膜环状核苷酸门控通道打开,促使细胞外Na+和Ca2+离子内流,Ca2+内流浓度升高后,促进细胞膜Cl−例子通道开放,促进Cl−外流,引起细胞去极化。在磷酸二酯酶作用下,促进cAMP水解,终止cAMP的作用(图1)[81]。此外,腺苷酸环化酶阻断剂和磷酸二酯酶抑制剂都可以调节嗅觉TAARs信号级联转导,而Golf敲除小鼠消除了嗅觉TAARs介导的嗅觉反应[11,46]。综上所述,内源性或外源性痕量胺通过TAAR/Golf/Adcy3/cAMP信号通路,将配体信号转化为电信号参与机体的各种生理过程,发挥重要的生理活性。
3. TAARs在脑神经系统中的作用
精神分裂症是一种行为异常的慢性精神疾病,病因尚未完全清晰,全基因组测序研究发现精神分裂症与TAARs基因突变存在较大关联[16]。Duan等[20]对包括大脑在内的各种人体组织的RT-PCR研究表明,TAAR4优先表达于涉及精神分裂症病理生理学的大脑区域。这些数据为TAAR4是精神分裂症的候选基因提供了强有力的初步证据。Berry等[19]发现TAAR5 mRNA在小鼠杏仁核、弓状核和下丘脑等脑区有表达。Espinoza等[49]对TAAR5基因敲除(TAAR5-KO)小鼠进行旷场试验、明暗过渡试验、高架迷宫试验等行为学测试,发现TAAR5-KO小鼠没有表现出明显的发育异常,但表现出较少的焦虑和抑郁样行为,另外TAAR5-KO小鼠脑区的5-HT及其代谢物的组织水平也显著下降。随后,Efimova等[82]发现TAAR5基因敲除小鼠呈现多巴胺神经元数量、纹状体中多巴胺及其代谢产物的水平以及GDNF mRNA水平均显著性升高,提示TAAR5基因敲除后多巴胺神经元增殖可能增加。大脑皮质γ节律参与的大脑区域之间的信息传递(交流)与认知功能障碍的发病机制紧密相关有关。Belov等[50]采用高效液相色谱法测定了小鼠纹状体中多巴胺及其代谢产物在给药TAAR5激动剂α-NETA后的变化,α-NETA可引起多巴胺能系统的显著调节,波动频率显著性增加,推测TAAR5激动剂α-NETA可以调节纹状体多巴胺的传递,并导致脑活动γ节律的显著改变,而γ节律的显著性变化与精神分裂症相关缺陷一致,这些观察结果表明,TAAR5在影响大脑病理的认知功能调节中发挥重要作用。自闭症谱系障碍是一种异质性的神经发育障碍,表现为不同程度的人际交流障碍、语言发育障碍、狭隘的兴趣和僵化的日常生活,并伴有整体认知功能分数的降低,Huang等[52]采用基因表达谱和加权基因共表达网络分析方法,研究发现在自闭症群体中TAAR7b和TAAR7h表达水平的显著性下调,提示TAARs和相关基因调控网络可能在自闭症中发挥重要作用。最近,通过转录组学和肽组学技术,TAARs(特别是TAAR6)的失调已经在以认知障碍为特征的大脑疾病中被发现[47]。上述研究提示TAARs在脑神经系统中多种疾病的调控发挥十分重要的作用,也将是未来研究神经精神类疾病的重要靶点。
尽管TAAR在脑区广泛分布,但脑神经系统中的TAARs的配体研究较少,这也成为限制TAARs作为在精神神经类疾病研究开发的一类重要靶点的弊端,食物中丰富的生物胺为TAARs配体的研究提供了广阔的空间。未来的研究可以了解食物中不同的生物胺与TAARs之间的相互作用,根据TAARs在疾病发生与发展的进程中扮演的作用,特异性修饰或改造成为TAARs激动剂或者拮抗剂,可能成为寻找新的药理活性化合物的新途径。
4. TAARs在免疫系统中的作用
TAARs在免疫系统中发挥重要的作用,其中TAAR1的研究相对较为广泛,例如,Panas等[15]在恒河猴B细胞系中检测到TAAR1蛋白的高水平表达,TAAR1受体表达的增加伴随淋巴细胞免疫激活,提示TAAR1可能在免疫反应的产生或调节中发挥作用,靶向TAAR1的药物也可能改变免疫功能。TAAR1广泛存在于正常或恶性淋巴瘤细胞中,而TAAR1激动剂可显著性引起恶性淋巴瘤细胞的凋亡,正常细胞敏感性较差,提示TAAR1激动剂对白血病、淋巴瘤以及更广泛的白细胞病变具有治疗潜力[83]。然而,随着近年来异位嗅觉TAARs的研究不断深入,异位嗅觉TAARs在免疫系统中的作用也逐渐被科学家们发现,例如,Babusyte等[23]对人的白细胞进行分离,在分离的5种白细胞(中性粒细胞、单核细胞、T细胞、B细胞和自然杀伤细胞)中,除TAAR8外,其余5种均有表达,提示TAARs可能在这些细胞中发挥作用。白细胞中TAAR1和TAAR2是免疫细胞功能激活和调节的重要媒介,参与细胞迁移、细胞因子和免疫球蛋白的产生等过程。Nelson等[84]在小鼠脾脏分离的小鼠B细胞和自然杀伤细胞中发现表达多种TAARs。
免疫应答是机体自我保护的一个重要调节机制,食物中的多种物质可引起由多种免疫细胞参与的免疫反应,TAARs参与多种免疫细胞的调节,但是未见有明确研究提示食物中的特定生物胺引起的免疫反应是由TAARs介导,因此,将来的研究也可将这种可能性纳入研究的考量范围。
5. TAARs在内分泌系统中的作用
与健康人群相比,TAAR1的单核苷酸突变频率在胰岛素分泌受损的肥胖和超重患者中更高[85]。TAAR1选择性小分子激动剂显著性增加INS1E细胞和人胰岛中葡萄糖依赖性胰岛素的分泌,提高小鼠血浆酪酪肽(Peptide YY, PYY)和胰高血糖素样肽-1(Glucagon-like peptide 1, GLP-1)水平[86]。Illumina HiSeq基因表达分析显示,在高糖和脂肪酸的作用下,TAARs的表达显著性下调,异戊胺(TAAR3激动剂)、酪胺(TAAR1和TAAR4激动剂)或2-苯乙胺(TAAR1和TAAR4激动剂)使INS-1细胞cAMP水平显著升高,胰岛素分泌增强,提示嗅觉TAARs可能在β细胞中发挥调节胰岛素分泌的作用[14]。糖尿病是一组由多病因引起的以慢性高血糖为特征的代谢性疾病,其中最常见的并发症之一是糖尿病肾病,Ruiz-Hernández等[87]研究发现TAAR3、TAAR5和TAAR9在糖尿病大鼠肾脏中的表达显著性减少,提示TAAR3、TAAR5和TAAR9在肾脏中参与肾脏的保护作用,虽确切生理作用现有研究还未明确,但可作为改善糖尿病性肾病研究中的有效靶点。TAARs多态性或饮食变化导致的生物胺供应增加可能会破坏痕量胺能稳态,同时微生物更有效地粘附和浸润肠上皮,导致功能性微生物失调。
6. TAARs在心血管系统的作用
三碘甲状腺原氨酸是由甲状腺滤泡细胞合成及分泌的激素,研究提示三碘甲状腺原氨酸作为一种新的信号分子可以快速影响甲状腺激素作用的几个生理表现,例如,体温、心率和心输出量。与甲状腺激素不同,研究人员猜测三碘甲状腺原氨酸可能通过TAAR1或其他亚型的TAARs发挥生理功能。Chiellini等[37]研究发现大鼠心脏中除TAAR1表达外,TAAR2、TAAR3、TAAR4以及TAAR8a均有表达。Frascarelli等[88]研究发现β-苯乙胺、章胺和色胺会产生负性肌力效应,例如,发挥减少心输出量和主动脉压的作用。
心血管系统类疾病的发生率与人们日常生活的饮食息息相关,随着人们日常生活水平的提高,肥胖、高血压、高血脂、非酒精性脂肪肝等代谢性疾病的发病趋势持续走高。现有的研究提示多种生物胺在心血管系统发挥重要的生理作用,但是其发挥作用的效应靶点暂不明晰,TAARs是否是相关生物胺发挥作用的靶点将是未来重要的研究方向。
7. 总结与展望
GPCRs家族是人类中最庞大的膜蛋白家族,也是很多药物的重要靶点,据统计,靶向GPCR的药物销量占全球市场的40%[3]。GPCRs一直是药物研发的重要靶点之一,因为它们能够调控多种广泛的生理过程,而且在细胞表面具有可以成药的靶点。而且,最近对GPCRs的研究开辟了开发GPCR药物的新策略。例如,靶向GPCR的别构结合位点可以改变受体的结构、动态平衡和功能,从而提高治疗效果。而且,对受体激活机制的深入研究让研究人员可以设计出靶向激活特定细胞内信号通路的激动剂,从而减少因激活其它信号通路产生的副作用。
TAARs是一类新型的G蛋白偶联受体,参与人体的多种生理过程,除TAAR1近年来在精神神经疾病模型中的广泛研究,其他类型的受体也广受关注。由于TAARs的异位分布,其特殊的生理作用更加值得关注,所以人类TAAR超家族中一些成员可能是治疗精神和/或代谢障碍的靶点,但需要进一步的研究才能得出明确的结论。人体痕量胺的生物合成和储存,释放后的降解以及再摄取,都受到严格的调控,在许多病理条件下,痕量胺含量水平的不平衡是导致大脑神经系统功能的改变、免疫系统的失衡、能量代谢的紊乱的重要原因。食物中含有多种生物胺,人体摄入后也会在机体内发挥重要的生理活性。但是,为了充分理解痕量胺和TAAR家族的生理相关性,还有许多重要的问题需要解决。以下几个研究方向值得关注:
a. 异位嗅觉TAARs的组织分布及功能:现有研究关于TAARs在脑组织或其他组织分布的可用数据仍然较少,需要更详细的研究TAARs不同组织的分布情况。将来可通过原位杂交技术检测特定受体表达,以及通过对TAARs的靶向敲低/敲除进行表型特征的研究,来进一步阐明其生物学功能以及其作为潜在药物靶标的可行性上具有重要的作用。
b. 异位嗅觉TAARs的脱孤化研究:现有的研究提示仍有部分的TAARs为孤儿受体,体内内源性或外源天然配体仍未知,大多数TAAR缺乏明确作用的内源性配体,很大程度上减缓了该领域的进展。去孤儿化即为孤儿受体寻找同源配体,已经成为现代药物研发过程中一个重要的工作,例如,可利用高通量的方法分离出相应的天然配体等。食物中含有的多种胺类成分,为TAAR孤儿受体的配体高通量分子对接的筛选提供大量的化合物资源库。许多鉴别出的内源性TAAR配体都是简单的脱羧氨基酸,其他脱羧形式的氨基酸及其代谢产物可能是其他TAAR内源性配体的良好来源。
c. 活性生物胺的潜在靶点研究:目前大部分的注意力都集中在TAAR1及其在大脑中的作用上,异位TAAR在外周组织中的作用近几年也逐渐被发掘出来,研究TAARs在异位嗅觉组织调节中的作用将开启新的研究方向。人们日常饮食的食物中存在多种微量胺进入机体后发挥重要的药理活性,但是具体的作用靶点未知或暂不明晰,众多具有生理或药理作用的生物胺的作用靶点是否是TAARs值得研究。
d. 食物中的生物胺与TAARs之间的关系:食物中富含有多种生物胺,是TAARs外源配体研究的重要来源,随着TAAR得到更广泛的认识和其生物学效应的认知。食物中检测到的生物胺在充当TAARs配体时的安全有效剂量范围、发酵食品生产和质量控制中生物胺含量的限定、食物中不同生物胺的安全性评估等都值得在未来的研究中进一步探索。
e. 生物胺在食品领域的研究:值得注意的是,海洋鱼类产品、发酵产品、以及腌制食品等是食物中生物胺的主要来源。但是由于食物中由于微生物种类不同,其生长及产生物胺特性存在显著差异。研究不同产生物胺菌株的特性来采取针对性措施降低有害生物胺含量,以及开展食品生物胺的生物活性探索,例如,脑神经系统、内分泌系统、免疫系统等都存在TAARs的分布,食物中的生物胺作为外源配体,是如何通过TAARs发挥作用的等,都对食品的工业化生产具有重要意义。
-
表 1 TAARs及其内源性配体
Table 1 TAARs and their endogenous ligands
受体类型 种属 主要表达部位 检测方法 配体 参考文献 TAAR1 人、大鼠、小鼠 脑、脊髓、胰腺β细胞、胃、肠道、白细胞 RT-PCR β-苯乙胺、
p-酪胺[23,42-44] TAAR2 人、大鼠、小鼠 肠道、心脏、睾丸、白细胞 RT-PCR / [23,45-47] TAAR3 大鼠、小鼠 下丘脑、胰腺β细胞 RT-PCR 异戊胺 [48] TAAR4 大鼠、小鼠 大脑、胰腺β细胞 RT-PCR β-苯乙胺 [14] TAAR5 人、大鼠、小鼠 大脑、脊髓、肠道、睾丸、白细胞 RT-PCR 三甲胺 [49−50] TAAR6 人、大鼠、小鼠 大脑、肠道、睾丸、白细胞、肾脏 RT-PCR / [19,47,51] TAAR7a 大鼠、小鼠 大脑、肾脏 RT-PCR、
微阵列/ [52] TAAR7b 大鼠 大脑、胰腺β细胞 N,N-二甲基烷基胺 [14,52] TAAR7c 大鼠、小鼠 大脑、胰腺β细胞 / TAAR7d 大鼠、小鼠 大脑、胰腺β细胞 N,N-二甲基烷基胺 TAAR7e 大鼠、小鼠 大脑、胰腺β细胞 N,N-二甲基烷基胺 TAAR7f 小鼠 大脑 N,N-二甲基烷基胺 TAAR7g 大鼠 大脑、胰腺β细胞 / TAAR7h 大鼠 大脑、胰腺β细胞 N,N-二甲基烷基胺 TAAR8a 大鼠、小鼠 星形胶质细胞、嗅觉上皮、杏仁核、白细胞、
胃、肠、心脏、睾丸、肺、脾、肾、肌肉RT-PCR N-甲基哌啶 [46,53] TAAR8b 大鼠、小鼠 TAAR8c 大鼠、小鼠 TAAR9 人、大鼠、小鼠 嗅上皮、脊髓、肠、脾、骨骼肌、垂体、白细胞 RT-PCR N-甲基哌啶 [19,42]  下载: 导出CSV
下载: 导出CSV
表 2 食物中的生物胺及其受体
Table 2 Biogenic amines and their receptors in food
生物胺 受体类型 主要食物来源 参考文献 酪胺 TAAR1 牛奶以及奶制品(酪乳、牛奶、酸奶) [29,42,55,57,59] 芝士 肉类(鸡肝、猪肉、火腿) 鱼(鳕鱼、沙丁鱼、马鲛鱼、金枪鱼、虾) 水果(牛油果、香蕉、葡萄、柑橘、西瓜、覆盆子) 蔬菜(甜菜根、卷心菜、胡萝卜、花椰菜、黄瓜、莴苣、
甜红椒、大豆、菠菜、西红柿、土豆)β-苯乙胺 TAAR1
TAAR4奶酪、葡萄酒、巧克力 [42] 三甲胺 TAAR5 猪肉、鱼、虾 [69−70] 腐胺 / 肉类、腌制鱼类、腌制蔬菜 [66,68, 72] 尸胺 TAAR13c 肉类、腌制鱼类、腌制蔬菜 [66,68,72,77] 组胺 TAAR1 肉类、腌制鱼类、腌制蔬菜 [66,68,72,78] 亚精胺 / 猪肉、羊肉、牛肉、鸡肉 [71] 精胺 TAAR348 猪肉、羊肉、牛肉、鸡肉 [71,79] 色胺 TAAR1 烤肉、干发酵香肠 [80]
下载: 导出CSV
-
[1] Roth B L, Irwin J J, Shoichet B K. Discovery of new GPCR ligands to illuminate new biology[J]. Nat Chem Biol,2017,13(11):1143−1151. doi: 10.1038/nchembio.2490
[2] Wacker D, Stevens R C, Roth B L. How ligands illuminate GPCR molecular pharmacology[J]. Cell,2017,170(3):414−427. doi: 10.1016/j.cell.2017.07.009
[3] Hutchings C J, Koglin M, Olson W C, et al. Opportunities for therapeutic antibodies directed at G-protein-coupled receptors[J]. Nature Reviews Drug discovery,2017,16(9):787−810.
[4] Ihara S, Yoshikawa K, Touhara K. Chemosensory signals and their receptors in the olfactory neural system[J]. Neuroscience,2013,254:45−60. doi: 10.1016/j.neuroscience.2013.08.063
[5] Feldmesser E, Olender T, Khen M, et al. Widespread ectopic expression of olfactory receptor genes[J]. BMC Genomics,2006,7:121. doi: 10.1186/1471-2164-7-121
[6] Chen Z, Zhao H, Fu N, et al. The diversified function and potential therapy of ectopic olfactory receptors in non-olfactory tissues[J]. J Cell Physiol,2018,233(3):2104−2115. doi: 10.1002/jcp.25929
[7] Tong T, Ryu S E, Min Y, et al. Olfactory receptor 10J5 responding to α-cedrene regulates hepatic steatosis via the cAMP-PKA pathway[J]. Scientific Reports,2017,7(1):9471. doi: 10.1038/s41598-017-10379-x
[8] Tong T, Park J, Moon C, et al. Regulation of adipogenesis and thermogenesis through mouse olfactory receptor 23 stimulated by α-cedrene in 3T3-L1 cells[J]. Nutrients,2018,10(11):1781. doi: 10.3390/nu10111781
[9] Liberles S D, Buck L B. A second class of chemosensory receptors in the olfactory epithelium[J]. Nature,2006,442(7103):645−650. doi: 10.1038/nature05066
[10] Johnson M A, Tsai L, Roy D S, et al. Neurons expressing trace amine-associated receptors project to discrete glomeruli and constitute an olfactory subsystem[J]. Proc Natl Acad Sci USA,2012,109(33):13410−13415. doi: 10.1073/pnas.1206724109
[11] Zhang J, Pacifico R, Cawley D, et al. Ultrasensitive detection of amines by a trace amine-associated receptor[J]. J Neurosci,2013,33(7):3228−3239. doi: 10.1523/JNEUROSCI.4299-12.2013
[12] Borowsky B, Adham N, Jones K A, et al. Trace amines: Identification of a family of mammalian G protein-coupled receptors[J]. Proc Natl Acad Sci USA,2001,98(16):8966−8971. doi: 10.1073/pnas.151105198
[13] Bunzow J R, Sonders M S, Arttamangkul S, et al. Amphetamine, 3, 4-methylenedioxymethamphetamine, lysergic acid diethylamide, and metabolites of the catecholamine neurotransmitters are agonists of a rat trace amine receptor[J]. Mol Pharmacol,2001,60(6):1181−1188. doi: 10.1124/mol.60.6.1181
[14] Cripps M J, Bagnati M, Jones T A, et al. Identification of a subset of trace amine-associated receptors and ligands as potential modulators of insulin secretion[J]. Biochem Pharmacol,2020,171:113685. doi: 10.1016/j.bcp.2019.113685
[15] Panas M W, Xie Z, Panas H N, et al. Trace amine associated receptor 1 signaling in activated lymphocytes[J]. J Neuroimmune Pharmacol,2012,7(4):866−876. doi: 10.1007/s11481-011-9321-4
[16] John J, Kukshal P, Bhatia T, et al. Possible role of rare variants in trace amine associated receptor 1 in schizophrenia[J]. Schizophr Res,2017,189:190−195. doi: 10.1016/j.schres.2017.02.020
[17] Hussain A, Saraiva L R, Korsching S I. Positive darwinian selection and the birth of an olfactory receptor clade in teleosts[J]. Proc Natl Acad Sci USA,2009,106(11):4313−4318. doi: 10.1073/pnas.0803229106
[18] Lindemann L, Ebeling M, Kratochwil N A, et al. Trace amine-associated receptors form structurally and functionally distinct subfamilies of novel G protein-coupled receptors[J]. Genomics,2005,85(3):372−385. doi: 10.1016/j.ygeno.2004.11.010
[19] Berry M D, Gainetdinov R R, Hoener M C, et al. Pharmacology of human trace amine-associated receptors: Therapeutic opportunities and challenges[J]. Pharmacol Ther,2017,180:161−180. doi: 10.1016/j.pharmthera.2017.07.002
[20] Duan J, Martinez M, Sanders A R, et al. Polymorphisms in the trace amine receptor 4 (TRAR4) gene on chromosome 6q23.2 are associated with susceptibility to schizophrenia[J]. Am J Hum Genet,2004,75(4):624−638. doi: 10.1086/424887
[21] Chiellini G, Erba P, Carnicelli V, et al. Distribution of exogenous [125I]-3-iodothyronamine in mouse in vivo: Relationship with trace amine-associated receptors[J]. J Endocrinol,2012,213(3):223−230. doi: 10.1530/JOE-12-0055
[22] Ito J, Ito M, Nambu H, et al. Anatomical and histological profiling of orphan G-protein-coupled receptor expression in gastrointestinal tract of C57BL/6J mice[J]. Cell Tissue Res,2009,338(2):257−269. doi: 10.1007/s00441-009-0859-x
[23] Babusyte A, Kotthoff M, Fiedler J, et al. Biogenic amines activate blood leukocytes via trace amine-associated receptors TAAR1 and TAAR2[J]. J Leukoc Biol,2013,93(3):387−394. doi: 10.1189/jlb.0912433
[24] Dinter J, Mühlhaus J, Wienchol C L, et al. Inverse agonistic action of 3-iodothyronamine at the human trace amine-associated receptor 5[J]. PLoS One,2015,10(2):e0117774. doi: 10.1371/journal.pone.0117774
[25] Gozal E A, O'Neill B E, Sawchuk M A, et al. Anatomical and functional evidence for trace amines as unique modulators of locomotor function in the mammalian spinal cord[J]. Frontiers in Neural Circuits,2014,8:134.
[26] Vanti W B, Muglia P, Nguyen T, et al. Discovery of a null mutation in a human trace amine receptor gene[J]. Genomics,2003,82(5):531−536. doi: 10.1016/S0888-7543(03)00173-3
[27] Regard J B, Sato I T, Coughlin S R. Anatomical profiling of G protein-coupled receptor expression[J]. Cell,2008,135(3):561−571. doi: 10.1016/j.cell.2008.08.040
[28] Erspamer V. Active substances of the posterior salivary glands of octopus and the hypobranchial glands of the purpur snail[J]. Arzneimittelforschung,1952,2(6):253−258.
[29] Ghiretti F. Enteramine, octopamine, and tyramine in external and internal secretion of the posterior salivary gland in octopus[J]. Arch Sci Biol (Bologna),1953,37(5):435−441.
[30] Philips S R. Amphetamine, p-hydroxyamphetamine and beta-phenethylamine in mouse brain and urine after (-)- and (+)-deprenyl administration[J]. J Pharm Pharmacol,1981,33(11):739−741.
[31] Durden D A, Philips S R, Boulton A A. Identification and distribution of beta-phenylethylamine in the rat[J]. Can J Biochem,1973,51(7):995−1002. doi: 10.1139/o73-129
[32] Paterson I A, Juorio A V, Boulton A A. 2-Phenylethylamine: A modulator of catecholamine transmission in the mammalian central nervous system?[J]. J Neurochem,1990,55(6):1827−1837. doi: 10.1111/j.1471-4159.1990.tb05764.x
[33] Jones R S. Tryptamine: A neuromodulator or neurotransmitter in mammalian brain?[J]. Prog Neurobiol,1982,19(1-2):117−139. doi: 10.1016/0301-0082(82)90023-5
[34] Ibrahim K E, Couch M W, Williams C M, et al. Quantitative measurement of octopamines and synephrines in urine using capillary column gas chromatography negative ion chemical ionization mass spectrometry[J]. Anal Chem,1984,56(9):1695−1699. doi: 10.1021/ac00273a037
[35] Wang R, Wan L, Li Q, et al. Chemiluminescence of synephrine based on the cerium(IV)-rhodamine B system[J]. Luminescence,2007,22(2):140−146. doi: 10.1002/bio.937
[36] Scanlan T S, Suchland K L, Hart M E, et al. 3-Iodothyronamine is an endogenous and rapid-acting derivative of thyroid hormone[J]. Nat Med,2004,10(6):638−642. doi: 10.1038/nm1051
[37] Chiellini G, Frascarelli S, Ghelardoni S, et al. Cardiac effects of 3-iodothyronamine: A new aminergic system modulating cardiac function[J]. FASEB J,2007,21(7):1597−1608. doi: 10.1096/fj.06-7474com
[38] Khan M Z, Nawaz W. The emerging roles of human trace amines and human trace amine-associated receptors (hTAARs) in central nervous system[J]. Biomed Pharmacother,2016,83:439−449. doi: 10.1016/j.biopha.2016.07.002
[39] Broadley K J. The vascular effects of trace amines and amphetamines[J]. Pharmacol Ther,2010,125(3):363−375. doi: 10.1016/j.pharmthera.2009.11.005
[40] Da Silveira Agostini-Costa T. Bioactive compounds and health benefits of pereskioideae and cactoideae: A review[J]. Food Chem,2020,327:126961. doi: 10.1016/j.foodchem.2020.126961
[41] Reynolds G P, Gray D O. Gas chromatographic detection of N-methyl-2-phenylethylamine: A new component of human urine[J]. J Chromatogr,1978,145(1):137−140. doi: 10.1016/S0378-4347(00)81676-X
[42] Ohta H, Takebe Y, Murakami Y, et al. Tyramine and β-phenylethylamine, from fermented food products, as agonists for the human trace amine-associated receptor 1 (hTAAR1) in the stomach[J]. Biosci Biotechnol Biochem,2017,81(5):1002−1006. doi: 10.1080/09168451.2016.1274640
[43] Wolinsky T D, Swanson C J, Smith K E, et al. The trace amine 1 receptor knockout mouse: an animal model with relevance to schizophrenia[J]. Genes, Brain, and Behavior,2007,6(7):628−639. doi: 10.1111/j.1601-183X.2006.00292.x
[44] Moore C F, Valentina S, Pietro C. Trace amine-associated receptor 1 (TAAR1): A new drug target for psychiatry?[J]. Neurosci Biobehav Rev,2020,120(1):537−541.
[45] Bly M. Examination of the trace amine-associated receptor 2 (TAAR2)[J]. Schizophr Res,2005,80(2−3):367−368. doi: 10.1016/j.schres.2005.06.003
[46] Dewan A. Olfactory signaling via trace amine-associated receptors[J]. Cell Tissue Res,2020,383(1):395−407.
[47] Rutigliano G, Zucchi R. Molecular variants in human trace amine-associated receptors and their implications in mental and metabolic disorders[J]. Cell Mol Neurobiol,2020,40(2):239−255. doi: 10.1007/s10571-019-00743-y
[48] Li Q, Liberles S D. Odor sensing by trace amine-associated receptors[J]. Chemosensory Transduction,2016:67−80.
[49] Espinoza S, Sukhanov I, Efimova E V, et al. Trace amine-associated receptor 5 provides olfactory input into limbic brain areas and modulates emotional behaviors and serotonin transmission[J]. Front Mol Neurosci,2020,13:18. doi: 10.3389/fnmol.2020.00018
[50] Belov D R, Efimova E V, Fesenko Z S, et al. Putative trace-amine associated receptor 5 (TAAR5) agonist α-NETA increases electrocorticogram gamma-rhythm in freely moving rats[J]. Cell Mol Neurobiol,2020,40(2):203−213. doi: 10.1007/s10571-019-00716-1
[51] Pae C U, Yu H S, Amann D, et al. Association of the trace amine associated receptor 6 (TAAR6) gene with schizophrenia and bipolar disorder in a Korean case control sample[J]. J Psychiatr Res,2008,42(1):35−40. doi: 10.1016/j.jpsychires.2006.09.011
[52] Huang J Y, Tian Y, Wang H J, et al. Functional genomic analyses identify pathways dysregulated in animal model of autism[J]. CNS Neurosci Ther,2016,22(10):845−853. doi: 10.1111/cns.12582
[53] Mühlhaus J, Dinter J, Nürnberg D, et al. Analysis of human TAAR8 and murine Taar8b mediated signaling pathways and expression profile[J]. Int J Mol Sci,2014,15(11):20638−20655. doi: 10.3390/ijms151120638
[54] Tarján V, Jánossy G. The role of biogenic amines in foods[J]. Nahrung,1978,22(3):285−289. doi: 10.1002/food.19780220304
[55] Andersen G, Marcinek P, Sulzinger N, et al. Food sources and biomolecular targets of tyramine[J]. Nutr Rev,2019,77(2):107−115. doi: 10.1093/nutrit/nuy036
[56] Kaufmann A, Maden K. Easy and fast method for the determination of biogenic amines in fish and fish products with liquid chromatography coupled to orbitrap tandem mass spectrometry[J]. J AOAC Int,2018,101(2):336−341. doi: 10.5740/jaoacint.17-0407
[57] Ernest Frederick Gale. The production of amines by bacteria: The production of tyramine by Streptococcus faecalis[J]. Biochem J,1940,34(6):846. doi: 10.1042/bj0340846
[58] Bover-Cid S, Holzapfel W H. Improved screening procedure for biogenic amine production by lactic acid bacteria[J]. Int J Food Microbiol,1999,53(1):33−41. doi: 10.1016/S0168-1605(99)00152-X
[59] Moreno-Arribas V, Torlois S, Joyeux A, et al. Isolation, properties and behaviour of tyramine-producing lactic acid bacteria from wine[J]. J Appl Microbiol,2000,88(4):584−593. doi: 10.1046/j.1365-2672.2000.00997.x
[60] Ziegleder G, Stojacic E, Stumpf B. Occurrence of beta-phenylethylamine and its derivatives in cocoa and cocoa products[J]. Z Lebensm Unters Forsch,1992,195(3):235−238. doi: 10.1007/BF01202801
[61] Pastore P, Favaro G, Badocco D, et al. Determination of biogenic amines in chocolate by ion chromatographic separation and pulsed integrated amperometric detection with implemented wave-form at Au disposable electrode[J]. J Chromatogr A,2005,1098(1-2):111−115. doi: 10.1016/j.chroma.2005.08.065
[62] Hurst W J, Toomey P B. High-performance liquid chromatographic determination of four biogenic amines in chocolate[J]. Analyst,1981,106(1261):394−402. doi: 10.1039/an9810600394
[63] Bonetta S, Bonetta S, Carraro E, et al. Detection of biogenic amine producer bacteria in a typical Italian goat cheese[J]. J Food Prot,2008,71(1):205−209. doi: 10.4315/0362-028X-71.1.205
[64] Landete J M, Ferrer S, Polo L, et al. Biogenic amines in wines from three Spanish regions[J]. J Agric Food Chem,2005,53(4):1119−1124. doi: 10.1021/jf049340k
[65] García-Villar N, Saurina J, Hernández-Cassou S. High-performance liquid chromatographic determination of biogenic amines in wines with an experimental design optimization procedure[J]. Anal Chim Acta,2006,575(1):97−105. doi: 10.1016/j.aca.2006.05.074
[66] 戴莹, 宋海勇, 吴曦, 等. 肉制品中生物胺的形成、检测和控制研究进展[J]. 肉类研究,2020,34(11):89−97. [Dai Y, Song H Y, Wu X, et al. Recent progress in the formation, detection and control of biogenic amines in meat[J]. Meat Research,2020,34(11):89−97. [67] Fraqueza M J, Alfaia C M, Barreto A S. Biogenic amine formation in turkey meat under modified atmosphere packaging with extended shelf life: Index of freshness[J]. Poult Sci,2012,91(6):1465−1472. doi: 10.3382/ps.2011-01577
[68] 杨姗姗, 王晓雯, 林翠苹. 水产品中生物胺的研究进展[J]. 青岛农业大学学报(自然科学版),2021,38(1):65−73. [Yang S S, Wang X W, Lin C P. Research progress of biogenic aminie in aquatic produts[J]. Journal of Qingdao Agricultural University (Natural Science),2021,38(1):65−73. [69] 郭大钧, 万建荣, 刘俊荣. 用三甲胺特效电极对海产鱼虾作质量评定[J]. 水产学报,1989(3):248−253. [Guo D J, Wan J R, Liu J R. Use of a TMA-specific electrode in determining the quality of marine fish and shrimp[J]. Journal of Fisheries of China,1989(3):248−253. [70] 胡彩虹, 许梓荣. 气相色谱法测定猪肉、鱼和虾中三甲胺的含量[J]. 食品科学,2001(5):62−64. [Hu C H, Xu Z R. Determi-nation of trimethylamine in pork, fish and shrimp by gas chromatography[J]. Food Science,2001(5):62−64. doi: 10.3321/j.issn:1002-6630.2001.05.020 [71] 周明珠, 熊光权, 乔宇, 等. 鮰鱼冷藏过程中气味和新鲜度的变化及相关性[J]. 肉类研究,2020,34(3):68−74. [Zhou M Z, Xiong G Q, Qiao Y. Changes and correlation of odor and freshness of channel catfish during cold storage[J]. Meat Research,2020,34(3):68−74. [72] 李梅, 汪冬冬, 唐垚, 等. 中国市售酱腌菜中生物胺含量分析[J]. 食品与发酵工业,2020:1−10. [Li M, Wang D D, Tang Y, et al. Analysis of the biogenic amines content in differently processed pickles from the Chinese market[J]. Food and Fermentation Industries,2020:1−10. [73] Provensi G, Passani M B, Costa A, et al. Neuronal histamine and the memory of emotionally salient events[J]. Br J Pharmacol,2020,177(3):557−569. doi: 10.1111/bph.14476
[74] Worm J, Falkenberg K, Olesen J. Histamine and migraine revisited: Mechanisms and possible drug targets[J]. J Headache Pain,2019,20(1):30. doi: 10.1186/s10194-019-0984-1
[75] Zare D, Muhammad K, Bejo M H, et al. Determination of urocanic acid, a compound implicated in histamine toxicity, and assessment of biogenic amines relative to urocanic acid content in selected fish and fish products[J]. J Food Compost Anal,2015,37:95−103. doi: 10.1016/j.jfca.2014.06.014
[76] Mah J-H, Han H-K, Oh Y-J, et al. Biogenic amines in Jeotkals, Korean salted and fermented fish products[J]. Food Chem,2002,79(2):239−243. doi: 10.1016/S0308-8146(02)00150-4
[77] Dieris M, Ahuja G, Krishna V, et al. A single identified glomerulus in the zebrafish olfactory bulb carries the high-affinity response to death-associated odor cadaverine[J]. Scientific Reports,2017,7:40892. doi: 10.1038/srep40892
[78] Liberles SD. Trace amine-associated receptors: Ligands, neural circuits, and behaviors[J]. Curr Opin Neurobiol,2015,34:1−7.
[79] Scott A M, Zhang Z, Jia L, et al. Spermine in semen of male sea lamprey acts as a sex pheromone[J]. PLoS Biol,2019,17(7):e3000332. doi: 10.1371/journal.pbio.3000332
[80] Costa D J, Martínez A M, Ribeiro W F, et al. Determination of tryptamine in foods using square wave adsorptive stripping voltammetry[J]. Talanta,2016,154:134−140. doi: 10.1016/j.talanta.2016.03.063
[81] Reisert J, Zhao H. Perspectives on: Information and coding in mammalian sensory physiology: Response kinetics of olfactory receptor neurons and the implications in olfactory coding[J]. J Gen Physiol,2011,138(3):303−310. doi: 10.1085/jgp.201110645
[82] Efimova E V, Kozlova A A, Razenkova V, et al. Increased dopamine transmission and adult neurogenesis in trace amine-associated receptor 5 (TAAR5) knockout mice[J]. Neuropharmacology,2021,182:108373. doi: 10.1016/j.neuropharm.2020.108373
[83] Wasik A M, Millan M J, Scanlan T, et al. Evidence for functional trace amine associated receptor-1 in normal and malignant B cells[J]. Leuk Res,2012,36(2):245−249. doi: 10.1016/j.leukres.2011.10.002
[84] Nelson D A, Tolbert M D, Singh S J, et al. Expression of neuronal trace amine-associated receptor (Taar) mRNAs in leukocytes[J]. J Neuroimmunol,2007,192(1−2):21−30. doi: 10.1016/j.jneuroim.2007.08.006
[85] Mühlhaus J, Dinter J, Jyrch S, et al. Investigation of naturally occurring single-nucleotide variants in human TAAR1[J]. Front Pharmacol,2017,8:807. doi: 10.3389/fphar.2017.00807
[86] Raab S, Wang H, Uhles S, et al. Incretin-like effects of small molecule trace amine-associated receptor 1 agonists[J]. Molecular Metabolism,2016,5(1):47−56. doi: 10.1016/j.molmet.2015.09.015
[87] A R-H, Cabrera-Becerra S, Vera-Juárez G, et al. Diabetic nephropathy produces alterations in the tissue expression profile of the orphan receptors GPR149, GPR153, GPR176, TAAR3, TAAR5 and TAAR9 in wistar rats[J]. Nucleosides, Nucleotides & Nucleic Acids,2020,39(8):1150−1161.
[88] Frascarelli S, Ghelardoni S, Chiellini G, et al. Cardiac effects of trace amines: Pharmacological characterization of trace amine-associated receptors[J]. Eur J Pharmacol,2008,587(1−3):231−236. doi: 10.1016/j.ejphar.2008.03.055
 下载:
下载:

计量
- 文章访问数:
- HTML全文浏览量:
- PDF下载量:
