


Effects of Neohesperidin on the Differentiation of Adipocyte and the Underlying Mechanism in 3T3-L1 Preadipocytes
-
摘要: 为探讨新橙皮苷对脂肪细胞脂肪形成的影响及其作用机制,采用MTS法检测新橙皮苷对3T3-L1前脂肪细胞的细胞活力的影响,确定新橙皮苷的作用浓度后,油红O染色法和分光光度法测定3T3-L1脂肪细胞的分化及胞内甘油三酯的含量,RT-PCR测定脂肪形成相关基因CCAAT增强子结合蛋白α(CCAAT/enhancer binding proteinα,C/EBPα)和过氧化物酶体增殖激活受体γ(Peroxisome proliferators-activated receptorγ,PPARγ)mRNA表达,Western蛋白印迹法检测蛋白激酶B(Protein kinase B,PKB/Akt)、糖原合成酶激酶3β(Glycogen synthase kinase3β,GSK3β)和糖原合成酶(Glycogen synthase,GS)的磷酸化水平;利用GSK3β选择性的抑制剂LiCl作用3T3-L1脂肪细胞后,检测新橙皮苷对3T3-L1细胞分化、胞内甘油三酯含量、C/EBPα、PPARγ及aP2蛋白表达的影响。结果表明,新橙皮苷能极显著抑制脂肪细胞分化和胞内甘油三酯的形成(P<0.01),激活Akt信号通路,促进p-Akt和p-GSK3β水平增加,极显著抑制C/EBPα、PPARγ、aP2 mRNA和蛋白表达(P<0.01)。新橙皮苷的这些影响部分地被GSK3β抑制剂LiCl所抑制(P<0.01)。新橙皮苷能够通过激活Akt/GSK3β信号通路抑制脂肪细胞分化。
-
关键词:
- 新橙皮苷 /
- 蛋白激酶B/糖原合酶激酶-3β信号通路 /
- 脂肪细胞分化
Abstract: This investigation aimed to identify the effects of neohesperidin on the differentiation of adipocyte and the underlying mechanism in 3T3-L1 preadipocytes. The cell viabilities of neohesperidin were detected in 3T3-L1 cells by MTS assay. The intracellular accumulation of lipids was visualized with Oil-red O staining and spectrophotometry analysis. The mRNA of CCAAT/enhancer binding protein α(C/EBPα) and peroxisome proliferators-activated receptorγ(PPARγ), adipogenic-specific genes during adipocyte differentiation, were measured by RT-PCR. Glycogen synthase kinase3β (GSK3β), glycogen synthase (GS) andprotein phosphorylation, the Akt signaling pathway, were analysed by immunoblotting. To confirm this GSK3β effect, 3T3-L1 cells were incubated with neohesperidin and GSK3β inhibitor (LiCl), and the intracellular accumulation of lipids and the level of adipogenic-specific protein were measured.The results showed that, neohesperidin significantly inhibited adipocyte differentiation and intracellular triglyceride formation (P<0.01), activated Akt signaling pathway, and promoted p-Akt and p-gsk3β, significantly inhibited C/EBPα, PPARγ, the expression of AP2 mRNA and protein (P<0.01). These effects were partly reversed by inhibition of GSK3β activity by LiCl. In summary, neohesperidin suppresses adipocyte differentiation via the Akt/GSK3β signaling pathway in 3T3-L1 preadipocytes. -
通常肥胖引发的代谢综合征相关并发症的集合会显著增加心血管疾病和死亡的风险[1-3]。肥胖在细胞水平上的特征是脂肪组织中从前脂肪细胞分化出的脂肪细胞数量的增加和脂肪细胞体积的增大,而抑制前脂肪细胞向脂肪细胞分化可以减少脂肪组织的质量,从而减少肥胖的发生[4]。
脂肪细胞分化是一个复杂的过程。研究发现转录因子过氧化物酶体增殖激活受体家族(Peroxisome proliferators-activated receptors,PPARs)和CCAAT/增强子结合蛋白家族(CCAAT/enhancer binding proteins,C/EBPs)是调节脂肪细胞分化的主要转录因子[5],其中PPARγ具有脂肪组织特异性,通过与配体结合被激活后再与受体结合形成PPAR-RXR异二聚体,进一步同反应元件结合后激活靶基因,最终导致脂肪细胞分化成熟,而C/EBPα与PPARγ能互相激活,促进PPARγ高表达,保持分化细胞的表型。丝氨酸/苏氨酸蛋白激酶B(Protein kinase B,PKB/Akt)在脂肪细胞分化中发挥重要作用。胰岛素信号通过磷脂酰肌醇激酶(Phosphatidylinositol 3-kinase,PI3K)激活Akt,诱导下游靶点糖原合成酶激酶3β(Glycogen synthase kinase3β,GSK3β)的丝氨酸/苏氨酸磷酸化,从而磷酸化C/EBPβ、C/EBPα和糖原合成酶(Glycogen synthase,GS),进一步抑制脂肪细胞分化[6-7]。
新橙皮苷(Neohesperidin)是一种具有强烈苦味的黄酮类化合物,广泛存在于酸橙(Citrus aurantium)、甜橙(Citrus Sinensis)、枳实(Fructus aurantii immaturus)、枳壳(
Bitter Orange Fructus aurantii)等食药芸香科植物和茜草科植物中[8]。近年来越来越多的药理学研究发现,新橙皮苷能抑制心肌肥厚和重构[9],抑制过敏反应[10],减轻肺纤维化[11],能抗衰老,延长酿酒酵母寿命[12],抑制肿瘤发生发展[13-14],减少神经细胞凋亡[15],改善骨质疏松[16-17]等。另外在调节脂质方面,新橙皮苷在加入游离脂肪酸(free fatty acids,FFAs)的肝细胞中表现出较强的降血脂作用,并逆转急性或慢性血脂异常小鼠的脂质病变[18],新型新橙皮苷二氢查耳酮类似物还能通过Nrf2途径抑制人脂肪来源干细胞的成脂分化[19]。这些结果显示,新橙皮苷具有潜在的抗高血脂的活性。本研究在此基础之上,探讨新橙皮苷对脂肪细胞脂肪形成的影响及其作用机制,为新橙皮苷的减重/减脂机制提供了理论上的支撑,并为膳食补充剂的实际应用提供了科学的实验依据。 1. 材料与方法
1.1 材料与仪器
小鼠胚胎成纤维3T3-L1细胞株(前脂肪细胞) 中国科学院上海细胞库;新橙皮苷(纯度≧90%)、胰岛素、3-异丁基-1-甲基黄嘌呤(3-isobutyl-1-methylxanthine,IBMX)、地塞米松、LiCl(纯度≧99%) 美国Sigma‐Aldrich公司;CellTiter 96 AQueous单溶液细胞增殖检测试剂(MTS) 美国Promega公司;DMEM培养基 美国Hycolon公司;胎牛血清 以色列BI公司;甘油三酯检测试剂盒 北京普利莱基因技术有限公司;PPARγ单克隆抗体、C/EBPα单克隆抗体、磷酸化Akt(Ser473)单克隆抗体、磷酸化GSK3β (Ser9)单克隆抗体、GS单克隆抗体、磷酸化GS(Ser641)单克隆抗体 美国CST公司;TRIzol、逆转录试剂盒 美国Invitrogen公司;PCR试剂盒 TIANGEN公司。
HERAcell二氧化碳培养箱 150i美国Thermo Fisher公司;Infinite 200 PRO酶标仪 瑞士TECAN公司;Bio-RadChemiDoc XRS系统、 BIO-RAD CFX96荧光定量PCR 仪 美国Bio-Rad公司。
1.2 实验方法
1.2.1 细胞活力检测
3T3-L1前脂肪细胞于高糖DMEM培养基(含10%胎牛血清)中,5% CO2、37 ℃二氧化碳培养箱内培养24 h。取对数生长期的3T3-L1细胞(5×104个/mL)接入到96孔板中,细胞过夜培养约16~20 h,加入终浓度为1、10、50、100、200 μmol/L的新橙皮苷作用8 d后[4],根据CellTiter 96 AQueous单溶液细胞增殖检测试剂(MTS)说明书操作,每孔加入20 μL MTS试剂,然后在二氧化碳培养箱中继续培养1~4 h,最后于490 nm波长处读取吸光度值,测定细胞活力[20]。
1.2.2 3T3-L1前脂肪细胞诱导分化
根据实验方案,共分为5组,分别为对照组(正常DMEM培养基培养)、分化液组(含分化液的DMEM培养基培养)、罗格列酮组(含分化液的DMEM培养基+1.5 μmol/L罗格列酮培养)、新橙皮苷组(含分化液的DMEM培养基+1或10 μmol/L新橙皮苷培养)。
根据文献[21]报道,具体方法为取对数生长期的3T3-L1前脂肪细胞(5×105个/mL)接入到6孔板中,待细胞完全融合2 d后,换成分化液培养基(含5 μg/mL胰岛素、0.5 mmol/L IBMX和1 mmol/L地塞米松的DMEM培养基),在5% CO2、37 ℃下孵育2 d,然后换用只含有5 μg/mL胰岛素的DMEM培养基继续孵育2 d,随后再换用正常DMEM培养基继续培养,2 d换培养基1次,培养4 d,即共诱导分化8 d。在开始诱导细胞分化的同时,阳性药物罗格列酮(1.5 μmol/L)和不同浓度的新橙皮苷(1、10 μmol/L)被加入到分化培养基中,直到实验结束。
当考察Akt/GSK3β信号通路在新橙皮苷抑制3T3-L1前脂肪细胞分化中的作用时,在加入新橙皮苷的同时加入GSK3β抑制剂(20 mmol/L LiCl),其它培养步骤同上,共孵育8 d后再进行相关检测[22]。
1.2.3 油红O染色
经8 d诱导分化后,细胞用生理盐水轻柔地漂洗2次,空气中晾干,用4%甲醛固定1 h,然后加入0.6% (w/v)油红O染液(60%异丙醇和40%水配制)继续染色1 h,接着用60%异丙醇漂洗细胞3次,洗掉未着色的染料后,显微镜下拍照。拍照完毕后,用4%的NP-40溶解胞内被油红O染色的脂肪滴,充分溶解后,于490 nm波长处读取吸光度值。
1.2.4 甘油三酯含量测定
经8 d诱导分化后,细胞用冰生理盐水轻柔地漂洗2次,然后收集细胞,每孔加入200 μL生理盐水,50 W超声裂解1 min,在4 ℃转速为13000×g离心20 min,收集上清液。按照甘油三酯含量检测试剂盒操作说明检测。
1.2.5 RT-PCR检测mRNA表达
参照文献[23]报道,3T3-L1细胞诱导分化后,收集细胞,用Trizol提取细胞总RNA,取1 μg总RNA按照逆转录试剂盒说明进行逆转录,引物序列如下:PPARγ,forward 5ʹ‐TTTTCAAGGGTGCCAGTTTC-3ʹ,reverse 5ʹ-AATCCTT GGCCCTCTGAGAT-3ʹ;C/EBPα,forward 5ʹ-GAACAGCAACGAGTACCGGGTA-3ʹ,reverse 5ʹ-CCATGGCCTTGACCAAGGAG-3ʹ;aP2,forward 5ʹ-AACACCGAGATTTCCTTCAA-3ʹ,reverse 5ʹ-TCACGCCTTTCATAACACAT-3ʹ;β‐actin,forward 5ʹ-GACAACGGCTCCGGCATGTGCAAAG‐3ʹ,reverse 5ʹ-TTCACGGTTGGCCTTAGGGTTCAG‐3ʹ。循环参数为先在95 ℃时5 min,然后再按照以下条件进行35个循环,参数为95 ℃ 30 s,58 ℃ 30 s,72 ℃ 45 s,最后72 ℃延长5 min。
1.2.6 Western blot蛋白印迹检测蛋白表达
参照文献[24]报道,3T3-L1细胞诱导分化后,收集细胞,用pH为7.4的PBS漂洗2次,加入含蛋白酶抑制剂的裂解液,冰上裂解30 min,二喹啉甲酸法测定总蛋白质浓度,每孔上样总蛋白质量为10~20 μg,经SDS-PAGE凝胶电泳后,电转蛋白到PVDF膜,封闭液封闭后,加入各蛋白一抗抗体(Akt、p-Akt、PPARγ、C/EBPα、p-GS、GS、p-GSK3β、GSK3β和aP2),室温孵育2 h,漂洗3次后,再加入HRP标记的二抗,继续室温孵育2 h,漂洗3次,加入ECL化学发光底物显影,Quantity One软件分析蛋白印迹。
1.3 数据处理
统计分析使用Origin 8.0软件进行分析,组间比较采用ANOVA检验,实验数据均以
2. 结果与分析
2.1 新橙皮苷对3T3-L1前脂肪细胞活力的影响
如图1所示,不同浓度新橙皮苷(1、10、50、100、200 μmol/L)作用3T3-L1前脂肪细胞8 d后,与对照组(新橙皮苷浓度为0 μmol/L组)相比较,细胞活力均无显著性改变(P>0.05),提示本实验中直到新橙皮苷作用最大浓度为200 μmol/L时,脂肪细胞的完整性仍未被损伤。因此,在后续实验中,采用小于200 μmol/L的新橙皮苷进行脂肪细胞分化研究。
![]() 图 1 新橙皮苷对3T3-L1前脂肪细胞活力的影响(n=8)Figure 1. Effect of neohesperidin on 3T3-L1 cell viability (n=8)
图 1 新橙皮苷对3T3-L1前脂肪细胞活力的影响(n=8)Figure 1. Effect of neohesperidin on 3T3-L1 cell viability (n=8)2.2 新橙皮苷对3T3-L1前脂肪细胞诱导分化的影响
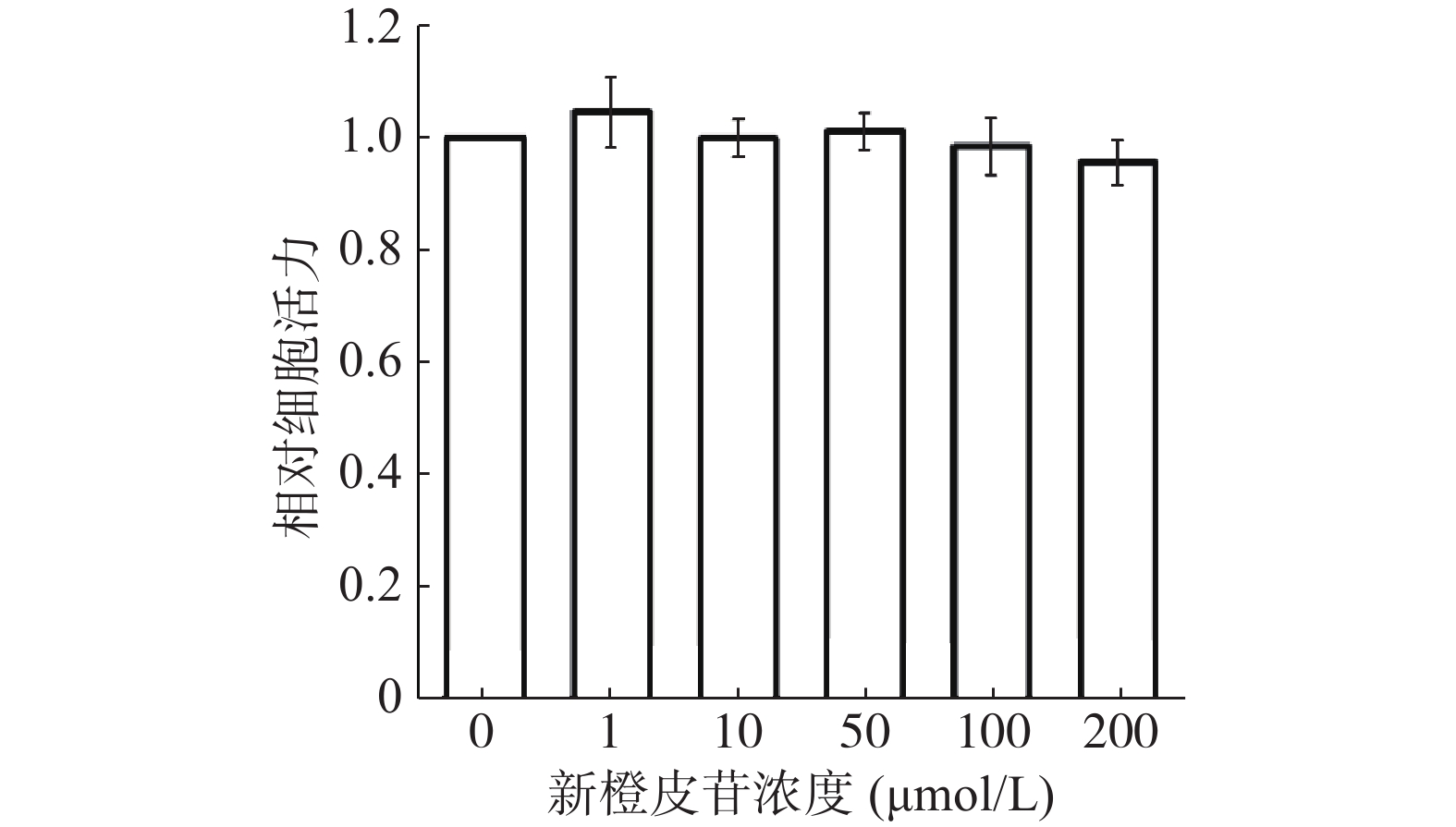
按照实验方法对3T3-L1前脂肪细胞进行8 d分化诱导,考察新橙皮苷在前脂肪细胞诱导分化中的影响。油红O染色结果如图2A所示,相对于无分化液组(对照组)比较,分化液组及阳性对照罗格列酮组的脂肪细胞变大,胞内脂滴含量明显增加。而加入了不同浓度(1和10 μmol/L)的新橙皮苷组脂滴含量较分化液组明显减少,脂肪细胞的分化呈剂量依赖性地明显降低。通过对分化细胞油红O染色后进行OD值检测,定量分析胞内脂肪的含量,如图2B所示,可以看出,与分化液组相比较,10 μmol/L新橙皮苷抑制分化率达到了25%。这些结果进一步通过胞内甘油三酯含量分析来证实,如图2C结果所示,与分化液组比较,10 μmol/L新橙皮苷组内甘油三酯的的含量减少了33%。提示新橙皮苷能抑制3T3-L1前脂肪细胞分化并抑制胞内脂质积累。
![]() 图 2 新橙皮苷对3T3-L1前脂肪细胞诱导分化的影响(n=5)注:A.油红O染色胞内脂滴(×400);B.胞内脂滴OD值;C.甘油三酯含量;*表示与分化液组相比差异显著(P<0.05),**表示与分化液组相比差异极显著(P<0.01)。Figure 2. Neohesperidin inhibits adipocyte differentiation of 3T3-L1 cells (n=5)
图 2 新橙皮苷对3T3-L1前脂肪细胞诱导分化的影响(n=5)注:A.油红O染色胞内脂滴(×400);B.胞内脂滴OD值;C.甘油三酯含量;*表示与分化液组相比差异显著(P<0.05),**表示与分化液组相比差异极显著(P<0.01)。Figure 2. Neohesperidin inhibits adipocyte differentiation of 3T3-L1 cells (n=5)2.3 新橙皮苷对诱导3T3-L1前脂肪细胞分化相关基因mRNA及蛋白质表达的影响
为了研究新橙皮苷对脂肪细胞分化相关基因mRNA及蛋白质表达的影响,本研究检测了Akt/GSK3β信号通路中Akt、GSK3β以及GS磷酸化水平以及下游转录因子PPARγ、C/EBPα和aP2 mRNA相对表达量变化。如图3A的RT-PCR分析显示,分化液组中3T3-L1前脂肪细胞完全分化后PPARγ、C/EBPα及aP2的mRNA表达增加,而新橙皮苷极显著降低了相应基因的表达(P<0.01),相对于分化液组,1和10 μmol/L新橙皮苷对PPARγ mRNA的表达分别降低了22%和69%,对C/EBPα mRNA的表达分别降低了15%和39%,同时对aP2 mRNA的表达分别降低了27%和44%。
![]() 图 3 新橙皮苷对诱导3T3-L1前脂肪细胞分化相关基因mRNA及蛋白质表达的影响注:A. PPARγ、C/EBPα和aP2 mRNA相对表达量;B. Western blot蛋白印迹图谱;C~E.分别为p-GS/GS、p-Akt/Akt和p-GSK3β/GSK3β的比例;**表示与分化液组相比差异极显著(P<0.01)。Figure 3. Effect of neohesperidin on the expression of mRNA and protein of adipogenic-specific genes during the differentiation of adipocyte
图 3 新橙皮苷对诱导3T3-L1前脂肪细胞分化相关基因mRNA及蛋白质表达的影响注:A. PPARγ、C/EBPα和aP2 mRNA相对表达量;B. Western blot蛋白印迹图谱;C~E.分别为p-GS/GS、p-Akt/Akt和p-GSK3β/GSK3β的比例;**表示与分化液组相比差异极显著(P<0.01)。Figure 3. Effect of neohesperidin on the expression of mRNA and protein of adipogenic-specific genes during the differentiation of adipocyteAkt信号通路,在胰岛素信号通路中,尤其是在脂肪细胞的分化过程中起到非常重要的作用[25]。为了阐明对新橙皮苷的作用及其抑制脂肪细胞分化的机制,通过western blotting方法检测了Akt及其底物GSK3β和GS的磷酸化表达水平。从图3B~图3E中可以看出,新橙皮苷降低了GS的磷酸化水平,但增加了Akt和GSK3β的磷酸化,说明新橙皮苷激活了Akt通路,降低了GSK3β的活性。
2.4 Akt/GSK3β信号通路在新橙皮苷抑制3T3-L1前脂肪细胞分化中的作用
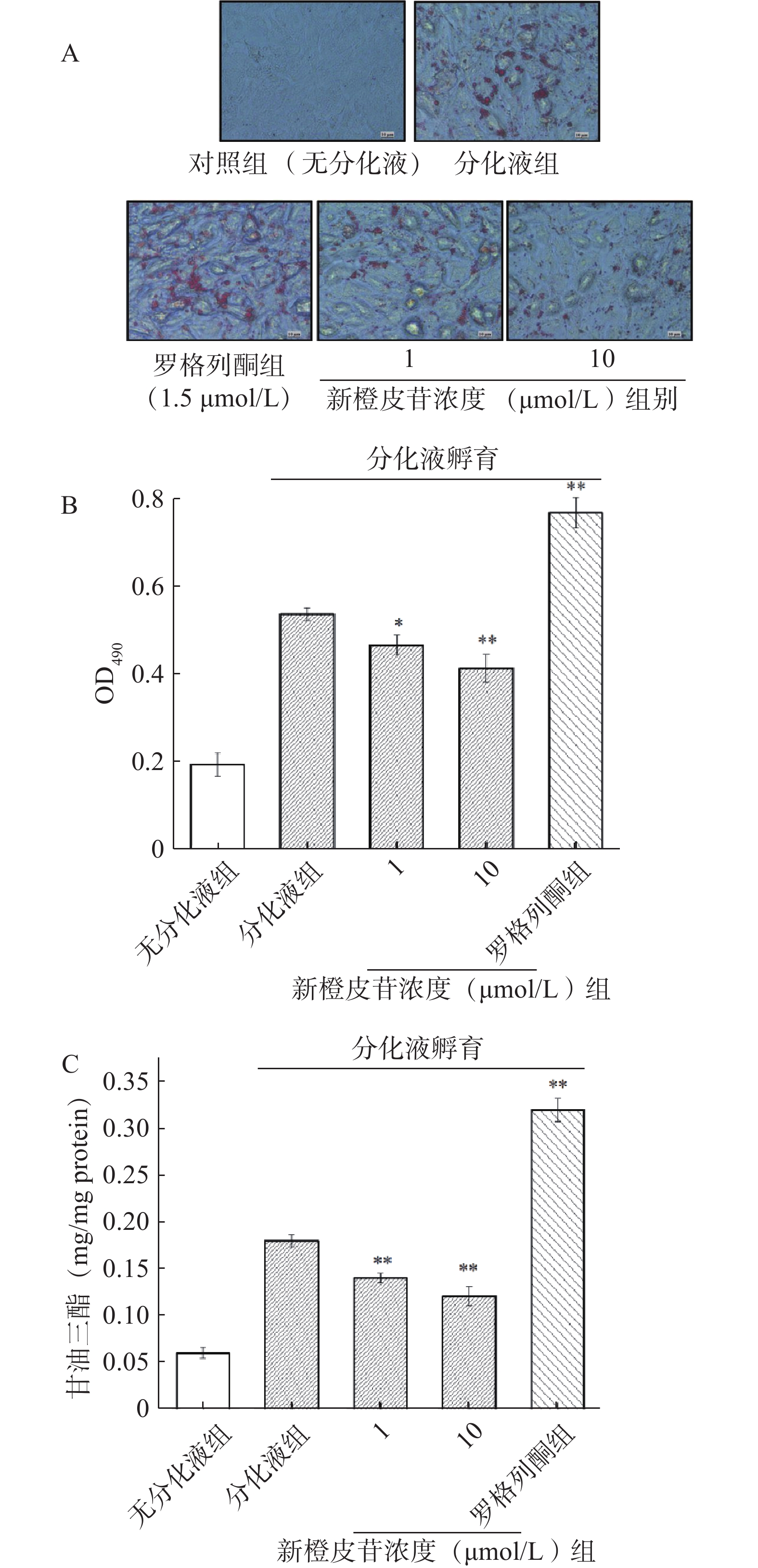
图3显示,新橙皮苷降低了脂肪分化相关蛋白PPARγ、C/EBPα以及aP2的mRNA的表达,也同时抑制了Akt信号通路中GSK3β的活性。据此,进一步探讨新橙皮苷抑制脂肪细胞分化和PPARγ表达是否与影响了GSK3β活性有关。为了证实GSK3β的作用,将3T3-L1细胞与新橙皮苷和GSK3β抑制剂(LiCl)共孵育8 d[22]。图4A~图4C显示,与LiCl共处理后,新橙皮苷介导的对脂肪细胞分化的抑制有拮抗作用。与这些结果一致的是,LiCl也减少了新橙皮苷诱导的脂肪分化相关蛋白表达水平的抑制作用,包括PPARγ、C/EBPα、aP2以及p-GS (图4D~图4H)。以上结果提示,新橙皮苷对脂肪细胞分化的抑制作用部分是通过抑制GSK3β蛋白活性来调控的。
![]() 图 4 Akt/GSK3β信号通路在新橙皮苷抑制3T3-L1前脂肪细胞分化中的作用(n=5)注:A.油红O染色胞内脂滴(×400);B.胞内脂滴OD值;C.甘油三酯含量;D. Western blot蛋白印迹图谱;E~H.分别为PPAR/β-Actin、C/EBPα/β-Actin、aP2/β-Actin和p-GS/GS的比例;**表示与分化液组相比差异极显著(P<0.01),##表示与新橙皮苷组相比差异极显著(P<0.01)。Figure 4. Akt/GSK3β is critical for the inhibition of neohesperidin-induced adipocyte differentiation (n=5)
图 4 Akt/GSK3β信号通路在新橙皮苷抑制3T3-L1前脂肪细胞分化中的作用(n=5)注:A.油红O染色胞内脂滴(×400);B.胞内脂滴OD值;C.甘油三酯含量;D. Western blot蛋白印迹图谱;E~H.分别为PPAR/β-Actin、C/EBPα/β-Actin、aP2/β-Actin和p-GS/GS的比例;**表示与分化液组相比差异极显著(P<0.01),##表示与新橙皮苷组相比差异极显著(P<0.01)。Figure 4. Akt/GSK3β is critical for the inhibition of neohesperidin-induced adipocyte differentiation (n=5)3. 讨论与结论
本研究结果表明,新橙皮苷能抑制3T3-L1前脂肪细胞分化并抑制胞内脂质积累,这种抑制作用可能是通过激活Akt/GSK3β信号通路从而抑制脂肪分化相关蛋白表达而实现的。
有研究表明,新橙皮苷有较好的调节脂质的能力,在加入游离脂肪酸(FFAs)的HepG2肝细胞中表现出较强的降血脂作用,逆转了由蛋黄诱导的急性或慢性血脂异常小鼠的脂质病变,明显改善肥胖模型DIO小鼠血浆、肝脏和腓肠肌的脂质含量,并显著减轻了肥胖小鼠的体重[18]。但是新橙皮苷对脂肪细胞脂肪形成的影响及其作用机制还不清楚。本研究首先在3T3-L1前脂肪细胞上检测了新橙皮苷对前脂肪细胞分化的影响,结果证实新橙皮苷能够部分抑制前脂肪细胞分化和胞内脂质积累,并且显著降低与脂肪细胞分化和脂肪积累密切相关的PPARγ和C/EBPα的mRNA的表达。另外,PPARγ能调节控制脂肪生成过程,上调下游脂肪生成和脂质代谢相关基因aP2[26]。研究了新橙皮苷对PPARγ的靶基因aP2的调节作用。新橙皮苷剂量依赖性下调分化的3T3-L1细胞中aP2 mRNA表达。由此可见,新橙皮苷在3T3-L1细胞中是通过抑制PPARγ表达,进而下调aP2的表达来抑制脂肪形成的。
Akt信号通路在调节脂肪形成中起重要作用[27-28]。GSK3β蛋白是Akt信号通路的下游蛋白,是参与细胞信号转导的关键蛋白。在Akt信号通路中,脂肪形成的调控是通过减少GSK3β蛋白磷酸化水平,增加GSK3β的活性,从而导致PPARγ、C/EBPα蛋白表达增加促进脂肪细胞分化[29]。有研究报道GSK3β蛋白激酶过度活跃与一些代谢紊乱有关,如2型糖尿病和肥胖[30]。结合这些文献报道及新橙皮苷对3T3-L1细胞中PPARγ表达的抑制作用,推测新橙皮苷通过靶向作用上游Akt/GSK3β及下游PPARγ信号通路来降低3T3-L1脂肪细胞的分化。结果发现,新橙皮苷级联增加了p-Akt (活性形式)和p-GSK3β (非活性形式)的含量,抑制了下游PPARγ、C/EBPα mRNA表达和p-GS的含量。这个结果提示Akt/GSK3β信号通路有可能参与了新橙皮苷对脂肪细胞分化的抑制作用。通过GSK3β特异性的抑制剂LiCl证实了这个假设,新橙皮苷对脂肪细胞分化的抑制作用被与LiCl共孵育得到部分逆转,而且LiCl也抑制了新橙皮苷降低脂肪分化相关蛋白的作用,包括PPARγ、C/EBPα、aP2和p-GS。因此,Akt/GSK3β信号通路在新橙皮苷抑制脂肪形成中起重要作用。
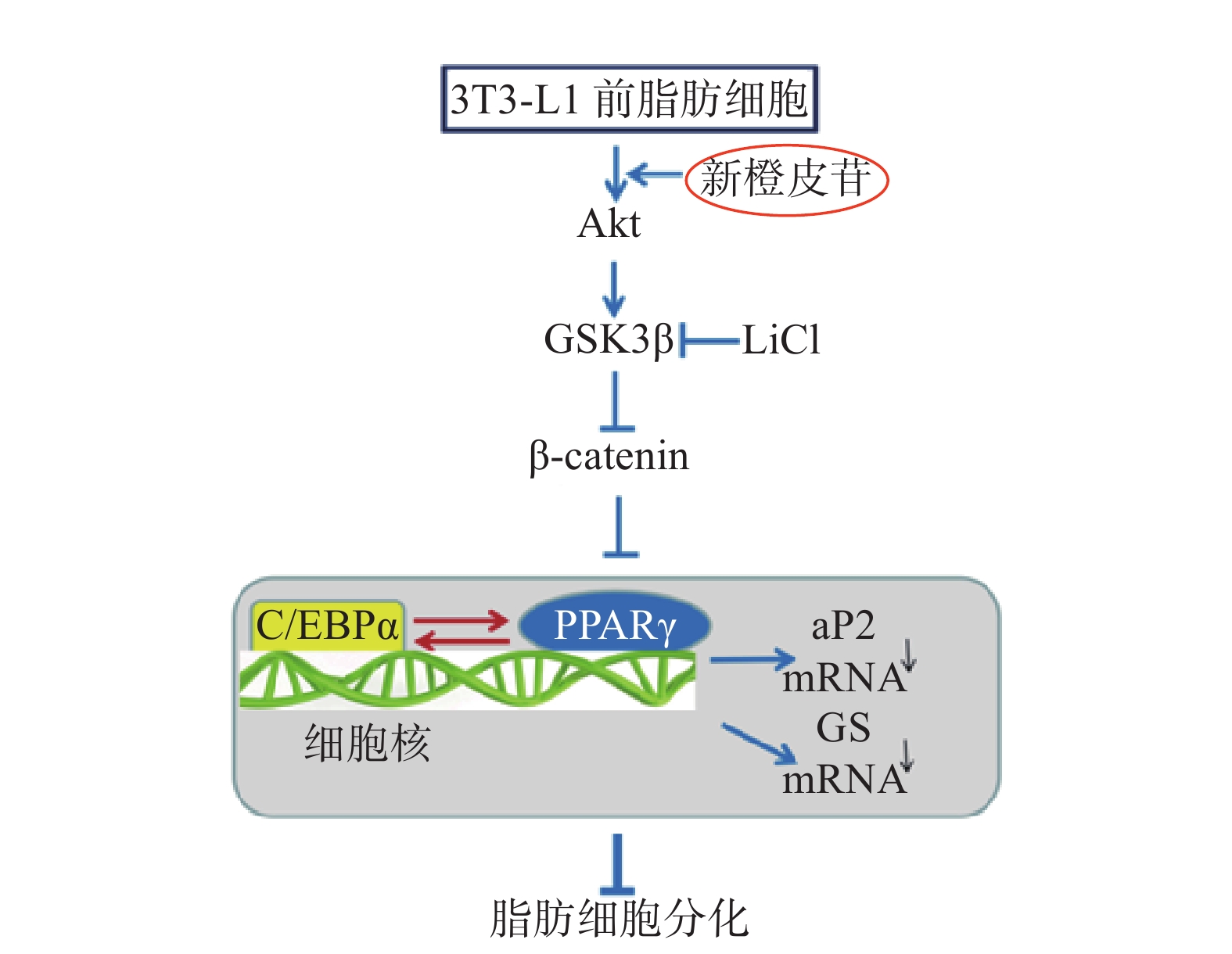
综上所述,新橙皮苷可通过激活Akt/GSK3β信号通路,增加GSK3β磷酸化水平,抑制其活力,进而减少下游脂肪分化相关蛋白PPARγ、C/EBPα、aP2的表达,减少脂肪细胞中脂质积累,抑制脂肪细胞分化(图5)。本实验在细胞水平初步评价了新橙皮苷的降脂效果,并探讨了其作用机制,为研究新橙皮苷在功能食品中的作用提供了一定的依据,但存在一定的局限性。进一步的实验可在动物水平研究其降脂减脂的效果以及在功能食品中的剂量设计,为新橙皮苷的研发提供数据支持。
![]() 图 5 新橙皮苷抑制3T3-L1前脂肪细胞分化细胞信号转导示意图Figure 5. Schematic diagram showing the inhibitory effect of neohesperidin on 3T3-L1 adipocyte differentiation
图 5 新橙皮苷抑制3T3-L1前脂肪细胞分化细胞信号转导示意图Figure 5. Schematic diagram showing the inhibitory effect of neohesperidin on 3T3-L1 adipocyte differentiation -
![]()
图 1 新橙皮苷对3T3-L1前脂肪细胞活力的影响(n=8)
Figure 1. Effect of neohesperidin on 3T3-L1 cell viability (n=8)
![]()
图 2 新橙皮苷对3T3-L1前脂肪细胞诱导分化的影响(n=5)
注:A.油红O染色胞内脂滴(×400);B.胞内脂滴OD值;C.甘油三酯含量;*表示与分化液组相比差异显著(P<0.05),**表示与分化液组相比差异极显著(P<0.01)。
Figure 2. Neohesperidin inhibits adipocyte differentiation of 3T3-L1 cells (n=5)
![]()
图 3 新橙皮苷对诱导3T3-L1前脂肪细胞分化相关基因mRNA及蛋白质表达的影响
注:A. PPARγ、C/EBPα和aP2 mRNA相对表达量;B. Western blot蛋白印迹图谱;C~E.分别为p-GS/GS、p-Akt/Akt和p-GSK3β/GSK3β的比例;**表示与分化液组相比差异极显著(P<0.01)。
Figure 3. Effect of neohesperidin on the expression of mRNA and protein of adipogenic-specific genes during the differentiation of adipocyte
![]()
图 4 Akt/GSK3β信号通路在新橙皮苷抑制3T3-L1前脂肪细胞分化中的作用(n=5)
注:A.油红O染色胞内脂滴(×400);B.胞内脂滴OD值;C.甘油三酯含量;D. Western blot蛋白印迹图谱;E~H.分别为PPAR/β-Actin、C/EBPα/β-Actin、aP2/β-Actin和p-GS/GS的比例;**表示与分化液组相比差异极显著(P<0.01),##表示与新橙皮苷组相比差异极显著(P<0.01)。
Figure 4. Akt/GSK3β is critical for the inhibition of neohesperidin-induced adipocyte differentiation (n=5)
-
[1] Elagizi A, Kachur S, Carbone S, et al. A Review of obesity, physical activity, and cardiovascular disease[J]. Current Obesity Reports,2020,9(4):571−581. doi: 10.1007/s13679-020-00403-z
[2] Dikaiou P, Bjorck L, Adiels M, et al. Obesity, overweight and risk for cardiovascular disease and mortality in young women[J]. European Journal of Preventive Cardiology,2020:2047487320908983. doi: 10.1177/2047487320908983
[3] Huang M Y, Wang M Y, Lin Y S, et al. The association between metabolically healthy obesity, cardiovascular disease, and all-cause mortality risk in Asia: Asystematic review and meta-analysis[J]. International Journal of Environmental Research and Public Health,2020,17(4):1320. doi: 10.3390/ijerph17041320
[4] Gao D, Zhang Y L, Yang F Q, et al. The flower of Edgeworthia gardneri (wall.) Meisn. suppresses adipogenesis through modulation of the AMPK pathway in 3T3-L1 adipocytes[J]. Journal of Ethnopharmacology,2016,191:379−386. doi: 10.1016/j.jep.2016.06.059
[5] Rosen E D, Hsu C H, Wang X, et al. C/EBPalpha induces adipogenesis through PPARgamma: A unified pathway[J]. Genes & Development,2002,16(1):22−26.
[6] Grimes C A, Jope R S. The multifaceted roles of GSK3beta in cellular signaling[J]. Progress in Neurobiology,2001,65:391−426. doi: 10.1016/S0301-0082(01)00011-9
[7] Ross S E, EricksonR L, Hemati N, et al. Glycogen synthase kinase 3 is an insulin-regulated C/EBPalpha kinase[J]. Molecular and Cellular Biology,1999,19:8433−8441. doi: 10.1128/MCB.19.12.8433
[8] Castillo J, Benavente O, Delrio J A. Naringin and neohesperidin levels during development of leaves, flower buds, and fruits of Citrus aurantium[J]. Plant Physiology,1992,99(1):67−73. doi: 10.1104/pp.99.1.67
[9] Zhang J, Fu X, Yang L, et al. Neohesperidin inhibits cardiac remodeling induced by Ang II in vivo and in vitro[J]. Biomedicine Pharmacotherapy,2020,129:110364. doi: 10.1016/j.biopha.2020.110364
[10] Zhao T, Hu S, Ma P, et al. Neohesperidin suppresses IgE-mediated anaphylactic reactions and mast cell activation via Lyn-PLC-Ca2+ pathway[J]. Phytotherapy Research,2019,33(8):2034−2043. doi: 10.1002/ptr.6385
[11] Guo J, Fang Y, Jiang F, et al. Neohesperidin inhibits TGF-beta1/Smad3 signaling and alleviates bleomycin-induced pulmonary fibrosis in mice[J]. European Journal of Pharmacology,2019,864:172712. doi: 10.1016/j.ejphar.2019.172712
[12] Guo C, Zhang H, Guan X, et al. The Anti-aging potential of neohesperidin and its synergistic effects with other citrus flavonoids in extending chronological lifespan of Saccharomyces cerevisiae BY4742[J]. Molecules,2019,24(22):4093. doi: 10.3390/molecules24224093
[13] Gong Y, Dong R, Gao X, et al. Neohesperidin prevents colorectal tumorigenesis by altering the gut microbiota[J]. Pharmacological Research,2019,148:104460. doi: 10.1016/j.phrs.2019.104460
[14] Xu F, Zang J, Chen D, et al. Neohesperidin induces cellular apoptosis in human breast adenocarcinoma MDA-MB-231 cells via activating the Bcl-2/Bax-mediated signaling pathway[J]. Natural Product Communications,2012,7(11):1475−1478.
[15] Wang J, Yuan Y, Zhang P, et al. Neohesperidin prevents Abeta25-35-induced apoptosis in primary cultured hippocampal neurons by blocking the S-nitrosylation of protein-disulphide isomerase[J]. Neurochemical Research,2018,43(9):1736−1744. doi: 10.1007/s11064-018-2589-5
[16] Tan Z, Cheng J, Liu Q, et al. Neohesperidin suppresses osteoclast differentiation, bone resorption and ovariectomised-induced osteoporosis in mice[J]. Molecularand Cellular Endocrinology,2017,439:369−378. doi: 10.1016/j.mce.2016.09.026
[17] 谭桢, 张丹, 程建文, 等. 新橙皮苷对破骨细胞分化影响的实验研究[J]. 微创医学,2018,13(6):719−722. [18] Wu H, Liu Y, Chen X, et al. Neohesperidin exerts lipid-regulating effects in vitro and in vivo via fibroblast growth factor 21 and AMP-activated protein kinase/sirtuin type 1/peroxisome proliferator-activated receptor gamma coactivator 1alpha signaling axis[J]. Pharmacology,2017,100(3-4):115−126. doi: 10.1159/000452492
[19] Han G E, Kang H T, Chung S, et al. Novel neohesperidin dihydrochalcone analogue inhibits adipogenic differentiation of human adipose-derived stem cells through the Nrf2 pathway[J]. International Journal of Molecular Sciences,2018,19(8):2215. doi: 10.3390/ijms19082215
[20] Chen M, Zhang J, Hu F, et al. Metformin affects the features of a human hepatocellular cell line (HepG2) by regulating macrophage polarization in a co-culture microenviroment[J]. Diabetes Metabolism Research Reviews,2015,31(8):781−789. doi: 10.1002/dmrr.2761
[21] Choi S S, Cha B Y, Lee Y S, et al. Magnolol enhances adipocyte differentiation and glucose uptake in 3T3-L1 cells[J]. Life Sciences,2009,84(25-26):908−914. doi: 10.1016/j.lfs.2009.04.001
[22] Zaragosi L E, Wdziekonski B, Fontaine C, et al. Effects of GSK3 inhibitors on in vitro expansion and differentiation of human adipose-derived stem cells into adipocytes[J]. BMC Molecular and Cell Biology, 2008, 9: 11.
[23] Lee S, Yang W K, Song J H, et al. Anti-obesity effects of 3-hydroxychromone derivative, a novel small-molecule inhibitor of glycogen synthase kinase-3[J]. Biochemical Pharmacology,2013,85(7):965−976. doi: 10.1016/j.bcp.2012.12.023
[24] 郭莉霞, 张永红, 殷钟意, 等. p-辛弗林通过激活AMPK-FoxO1信号通路抑制肝细胞葡萄糖生成[J]. 食品科学,2019,40(5):156−161. doi: 10.7506/spkx1002-6630-20171109-105 [25] Yun S J, Kim E K, Tucker D F, et al. Isoform-specific regulation of adipocyte differentiation by Akt/protein kinase Balpha[J]. Biochemical and Biophysical Research Communications,2008,371(1):138−143. doi: 10.1016/j.bbrc.2008.04.029
[26] Motade S A P, Richard A J, Hang H, et al. Transcriptional regulation of adipogenesis[J]. Comprehensive Physiology,2017,7(2):635−674.
[27] Sakaue H, Ogawa W, Matsumoto M, et al. Posttranscriptional control of adipocyte differentiation through activation of phosphoinositide 3-kinase[J]. Journalof Biological Chemistry,1998,273(44):28945−28952. doi: 10.1074/jbc.273.44.28945
[28] Xu J, Liao K. Protein kinase B/AKT 1 plays a pivotal role in insulin-like growth factor-1 receptor signaling induced 3T3-L1 adipocyte differentiation[J]. Journalof Biological Chemistry,2004,279(34):35914−35922. doi: 10.1074/jbc.M402297200
[29] Turk B E. Glycogen synthase kinase-3beta regulation: Another kinase gets in on the AKT[J]. FEBS Letters,2018,592(4):535−536. doi: 10.1002/1873-3468.12995
[30] Yen C L, Chao W C, Wu C H, et al. Phosphorylation of glycogen synthase kinase-3beta in metabolically abnormal obesity affects immune stimulation-induced cytokine production[J]. International Journal of Obesity,2015,39(2):270−278. doi: 10.1038/ijo.2014.93
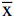
 下载:
下载:





 下载:
下载:



计量
- 文章访问数: 481
- HTML全文浏览量: 204
- PDF下载量: 29
