


Research Progress in Microbial Metabolic Engineering for Producing Monoterpene Aroma Products
-
摘要: 单萜及其衍生物是重要的植物天然产物,且具有多种生物学功能。该类物质在多个领域中均表现出较高的开发利用价值,目前已被作为优质香精香料广泛应用于食品、饮料、化妆品和医药工业中,市场需求日益增长。从植物中提取这些单萜芳香产品存在着来源少、含量低和分离困难等缺点,很难满足市场需求。因此,开发生产单萜芳香产品可再生的微生物资源来补充甚至代替原有的植物资源就具有重要的理论意义和应用价值。近年来,研究人员利用代谢工程技术已经成功构建了合成单萜芳香产品的微生物细胞工厂,达到了利用微生物合成法生产该类工业产品的目的。本文主要从菌株改造、发酵优化及产物分离等角度总结了相关产物合成的代谢工程实例,并分析了目前利用代谢工程改造微生物合成单萜芳香产品所面临的瓶颈问题及其可能的解决方法,旨在为构建异源、廉价、高效生产单萜芳香产品的微生物细胞工厂并最终实现其绿色制造提供参考。Abstract: Monoterpenes are an important class of plant natural products that exhibit a broad range of biological activities. Some monoterpenes and related derivatives have shown great potential for various industrial applications (e.g., one of the world’s most commonly used flavour and fragrance additives in foods, medicines and cosmetics), which lead to an increasing market demand for them. However, the low abundance or yield of most monoterpenes and derivatives in plants renders their isolation from plant sources non-economically viable. To meet the rapidly rising market demand for monoterpenes and derivatives, producing them by engineering microbial cells into microbial factories is becoming an attractive alternative approach which can overcome the aforementioned bottleneck, making it more sustainable and environmentally friendly. In recent years, these value-added products have successfully been obtained using metabolically engineered microbes. Here the latest examples of biosynthesis of monoterpene aroma products in the engineered microbes are reviewed . Furthermore, the current bottleneck issues and potential solutions are discussed. It is believed that the information provided here will make a significant contribution to further advancement of the microbial production of monoterpene aroma products through metabolic engineering.
-
Keywords:
- metabolic engineering /
- monoterpene /
- monoterpene derivatives /
- microbial cell factory /
- chassis
-
从代谢工程概念的首次提出到现在的30年左右的时间里,在分子生物学、基因组学、生物化学和基因工程等相关学科和技术的推动下,经过几代人的努力,目前代谢工程已经形成了一套比较完备的理论体系和技术方法。现阶段,代谢工程技术的基本路线就是首先利用先进的生物理论和技术对细胞的代谢途径及调控网络进行分析并提出合理的设计策略,再结合基因重组技术对相关途径和网络进行修饰、改造、扩展或者引入,从而实现改变细胞特性或者提高特定代谢产物产量的目的。
近年来,越来越多的科研人员投身于利用代谢工程技术构建微生物细胞工厂的研究工作。所谓的微生物细胞工厂如同一般意义上的工厂一样[1],由微生物细胞作为制造产品的生产厂房,代谢通路担任生产线,培养基提供生产原料和动力来源,而细胞内复杂的反馈和调控机制则是生产管理系统,工厂内各个部门之间密切配合,最终目的是实现目标产品产量的最大化。这其中最热门的研究就是利用代谢工程技术在不同微生物底盘中成功构建了包括单萜及其衍生物在内的多种植物天然产物的生物合成途径,如柠檬烯、紫苏醇、香叶醇、芳樟醇等。
单萜类植物天然产物是一类从植物体内分离出来的次生代谢产物,由两个异戊二烯骨架结构单位组成[2],具有丰富的药理学或生物学活性,广泛应用于食品、饮料、化妆品和医药工业中,其市场需求也日益增长。例如,柠檬烯就是一种典型的天然活性单萜物质,分子式为C10H16,属于单环单萜。它很早就被国际权威组织认定为“公认安全”(generally regarded as safe,GRAS)化合物,作为前体物可以转化合成多种高附加值的药物和香精化学品[3-4]。氢化后的柠檬烯由于凝固点较低而且不溶于水,在低温条件下作为添加剂能够有效改善燃料的性能[5],因此在能源领域也有很大的发展潜力。作为柠檬烯衍生物的紫苏醇、香叶醇是很好的香精原料,目前已被广泛应用于配制食品和日用产品;除此之外,这两种物质都有着非常出色的药理功效,如紫苏醇入药可以抑制癌细胞的迁移,逆转人体内已形成的肿瘤[6],而香叶醇作为抗炎剂在治疗炎症方面疗效显著[7]。另一种柠檬烯衍生物芳樟醇几乎每年都居于香料消费名单的榜首,实验表明芳樟醇也具有抑制细菌、真菌、病毒等微生物的活性[8-9],并且还具备缓解压力、提高睡眠质量等功效[10]。
迄今为止,人们获取活性植物天然产物的主要方法是植物提取分离法和有机合成法[11],但植物提取分离法存在植物生长周期长、产物提取难度大和效率低等问题,而化学合成法也存在着严重依赖化石燃料、反应效率低以及极易造成环境污染等缺点。近年来,随着代谢工程等相关技术取得了一系列的突破性发展,利用微生物细胞工厂异源合成植物天然产物有望成为替代植物提取分离法和化学合成法的一种有效策略。从环保和生态的角度来看,微生物合成法具有节约成本、减少环境污染等优势,因此是建设节约型社会,实现可持续发展的绿色生产方式,发展前景广阔。本文综述了近年来在不同的微生物底盘细胞中利用代谢工程技术构建细胞工厂从而生物合成一些单萜类植物天然产物方面的研究进展,并讨论了该方法目前所面临的瓶颈问题及其可能的解决措施,以期为在微生物细胞工厂中实现单萜类化合物的规模化、效益化工业生产提供可能的理论依据。
1. 在微生物底盘中所构建的单萜化合物异源合成途径
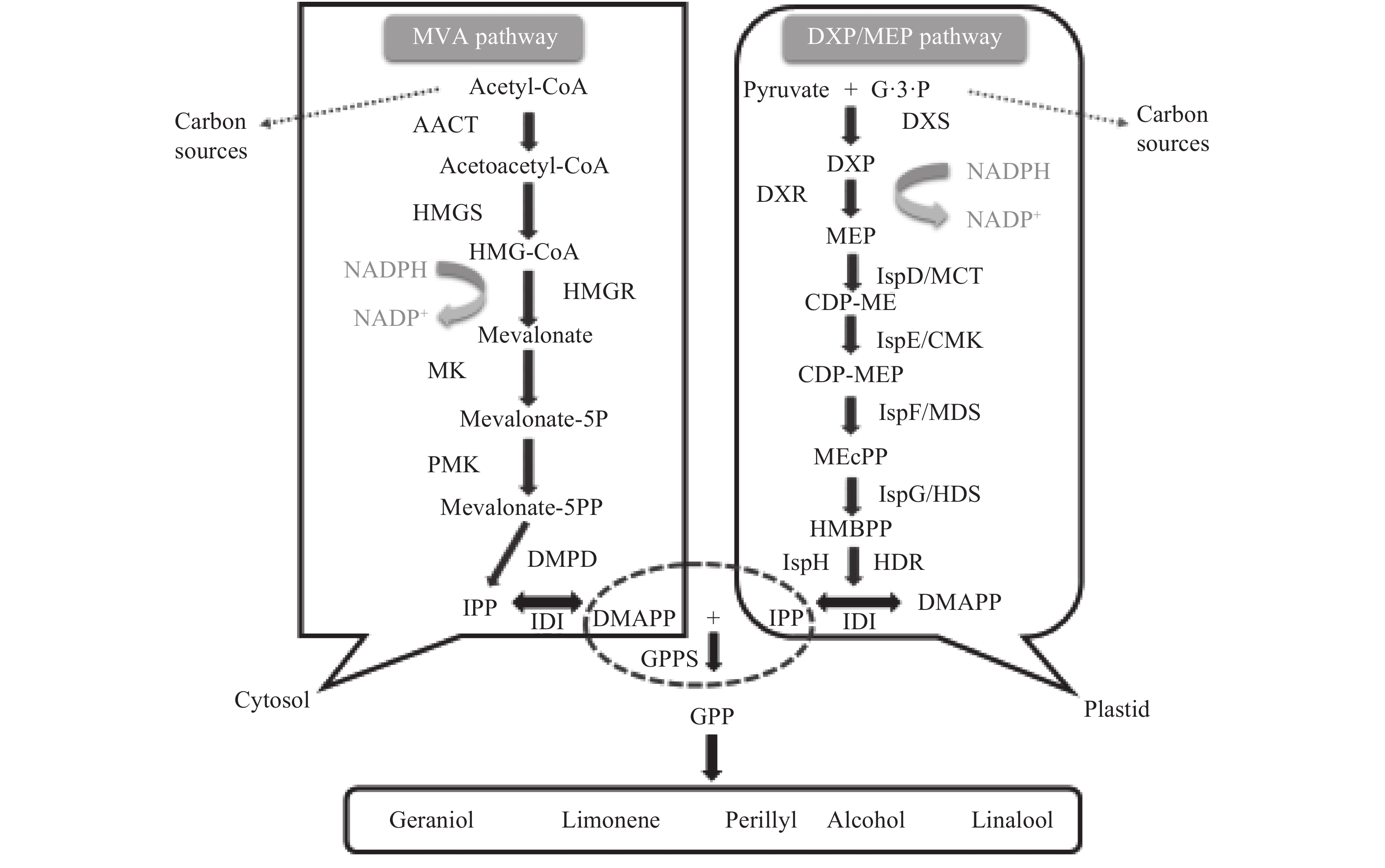
大多数微生物细胞本身并不具备合成单萜类植物天然产物的能力,研究人员利用代谢工程技术手段通过引入部分或完整代谢途径的方法重构了微生物的代谢网络,在不同微生物底盘中成功构建了单萜类化合物的异源合成途径(图1)。为了便于研究,大多数研究者会把单萜类化合物在微生物底盘中的合成过程分成三个模块:首先,由自然界中两种代谢路径合成植物单萜的共同前体物异戊烯基焦磷酸(isopentenyl pyrophosphate,IPP)和二甲基烯丙基焦磷酸(dimethylallyl pyrophosphate,DMAPP);随后,在单萜生物合成途径中关键酶——香叶基二磷酸合酶(geranyl diphosphate synthase,GPPS)的催化下,1分子IPP和1分子DMAPP合成直接前体香叶基焦磷酸(geranyl diphosphate,GPP);最后,GPP在一系列单萜合酶作用下生成各种单萜类化合物[11]。其中,IPP和DMAPP互为同分异构体,合成这两种物质的天然生物合成途径有两种,分别为甲羟戊酸(mevalonate,MVA)途径和甲基赤藓醇磷酸(methylerythritol phosphate,MEP)途径。MVA途径和MEP途径在合成场所、合成原料和途径酶等方面均存在明显差异:MEP途径主要存在于原核生物和植物的质体中,它以丙酮酸(pyruvate)和3-磷酸甘油醛(glyceraldehyde-3-phosphate,G3P)为合成原料,关键限速途径酶包括1-脱氧-D-木酮糖-5-磷酸合酶(1-deoxy-D-xylulose 5-phosphate synthase,DXS)、1-脱氧-D-木酮糖-5-磷酸还原异构酶(1-deoxy-D-xylulose-5-phosphate reductoisomerase,DXR)、4-羟基-3-甲基丁-2-烯基二磷酸合酶(4-hydroxy-3-methylbut-2-enyl diphosphate synthase,HDS)和4-羟基-3-甲基丁-2-烯基二磷酸还原酶(4-hydroxy-3-methylbut-2-enyl diphosphate reductase,HDR)等[12-13];MVA途径则主要存在于真核生物细胞质中,它以乙酰辅酶A(acetyl-CoA)为合成原料,关键限速酶为乙酰乙酰辅酶A硫解酶(acetoacetyl-CoA thiolase,AACT)、羟甲基戊二酰乙酰辅酶A合酶(hydroxymethylglutaryl-CoA synthase,HMGS)、羟甲基戊二酰辅酶A还原酶(hydroxymethylglutaryl-CoA reductase,HMGR)、甲羟戊酸激酶(mevalonate kinase,MK)和磷酸甲羟戊酸激酶(phosphomevalonate kinase,PMK)等[14]。尽管这两种代谢途径的合成逻辑完全不同,但是合成过程中却都产生了IPP、DMAPP、GPP这样的末端产物,并且都是通过中间产物交换和反馈调节作用来对微生物代谢网络进行调控,甚至在合成某些代谢物时彼此之间还会发生协同作用[15]。这些发现极大地激发了研究人员的探索兴趣,一方面驱使研究者去探寻是否在自然界中存在一条甚至多条有着更高生产效率的单萜类产物合成途径,另一方面启迪研究者通过合成生物学的手段去构建全新的非天然代谢途径来合成单萜类产物,为未来彻底解除限制目的产品高产的代谢瓶颈提供了更多的可能。
![]() 图 1 微生物单萜及其衍生物的生物合成途径Figure 1. Overview of the biosynthetic pathway of monoterpenes and their derivatives in microorganisms
图 1 微生物单萜及其衍生物的生物合成途径Figure 1. Overview of the biosynthetic pathway of monoterpenes and their derivatives in microorganisms2. 代谢工程改造微生物合成单萜芳香产品的策略及应用实例
2.1 代谢工程改造思路
近年来,通过利用代谢工程技术对特定微生物的代谢途径进行改造,研究者们已经成功实现了多种单萜芳香产品的微生物异源合成(表1)。代谢工程中的具体调控方式有很多种,大致可分为转录水平、翻译水平和蛋白质水平上的调控等。经过一代又一代科研工作者的努力,目前代谢工程改造过程中所使用到的技术和方法已经取得了很大的进步和突破。但目前在利用微生物合成单萜芳香产品的研究工作中,主要采用的还是强化目标产物合成途径中代谢通量的思路来对微生物底盘细胞进行改造,以实现目标产物产量的提高(表1),具体改造思路总结如下:
表 1 微生物单萜及其衍生物的产量和相关的途径改造策略Table 1. The production of monoterpenes and the related engineering strategies in microorganisms宿主 产品 最高产量(mg/L) 改造策略和发酵方式 参考文献 解脂耶氏酵母 柠檬烯 165.3 1)增加tNDPS1和tLS基因拷贝;
2)发酵条件优化;
3)分批补料发酵。[22] 解脂耶氏酵母 柠檬烯 D型11.705 1)单基因过表达基因HMGR;
2)发酵条件优化;
3)厨房废油作为单一碳源;
4)摇瓶培养。[17] L型11.088 大肠杆菌 柠檬烯 L型435 1)单质粒异源表达多个基因;
2)提高酶MK和PMK的含量;
3)表达外源基因HMGS和HMG1和截断型GPPS酶;
4)摇瓶培养。[20] 酿酒酵母 柠檬烯 D型0.12 1)戊烷萃取;
2)固相微萃取;
3)十二烷覆盖;
4)顶空捕捉。[23] L型0.49 酿酒酵母 香叶醇 1680 1)过表达基因tHMG1和IDI1;
2)利用计算机结构分析和建模来截短CrGES氮端的转运肽;
3)将GES和Erg20偶联为融合蛋白表达;
4)分批补料发酵。[2] 大肠杆菌 香叶醇 2000 1)添加十四酸异丙酯形成水-有机两相培养体系;
2)过表达乙酰酯酶Aes基因,验证了大肠杆菌能够水解乙酸香叶酯;
3)采用葡萄糖饥饿策略;
4)摇瓶培养。[21] 大肠杆菌 紫苏醇 105 1)诱导剂IPTG和阿拉伯糖诱导细胞来提高P450表达水平;
2)Amberlite树脂回收产品;
3)摇瓶培养。[20] 解脂耶氏酵母 芳樟醇 6.96 1)共过表达HMG1、IDI1、ERG20F88W-N119W三个基因;
2)添加混合碳源、辅助碳源来优化培养基;
3)摇瓶培养。[16] 酿酒酵母 芳樟醇 23.45 1)突变基因ERG20的K197位点;
2)线粒体区室化;
3)添加辅助碳源来优化培养基;
4)分批发酵。[18] 2.1.1 过表达关键酶基因
目前的研究结果基本能够确定单萜类产物合成的前体物GPP就是MVA途径的主要限速瓶颈,因此实现前体底物GPP的充足供应是提高单萜芳香产品产量的关键,而过表达MVA途径中的关键酶基因则是提高合成途径代谢通量的重要手段和方法。
羟甲基戊二酰辅酶A还原酶(HMGR)是MVA途径中的关键限速酶之一,其催化HMG-CoA生成甲羟戊酸的生化反应的强弱是决定微生物细胞内GPP产量的重要因素之一。华东理工大学花强团队[16]首先在解脂耶氏酵母产芳樟醇工程菌株中单基因过表达HMGR的编码基因HMG1,芳樟醇的产量与初始菌株相比显著增加,达到了0.52 mg/L。为了进一步促进产物的合成,该团队研究人员紧接着探讨了HMG1基因与MVA途径中磷酸甲羟戊酸激酶基因ERG8、乙酰乙酰辅酶A硫解酶基因ERG10、甲羟戊酸激酶基因ERG12或甲羟戊酸二磷酸脱羧酶基因ERG19组合表达时对芳樟醇产量的影响,发现与单独过表达HMG1的菌株相比,各基因组合过表达菌株合成芳樟醇的产量有了进一步的增加,其中共过表达HMG1和ERG12的工程菌株合成芳樟醇的能力最强,产量提升至0.84 mg/L。
由IDI1基因编码的异戊烯基二磷酸异构酶能够催化IPP异构化为DMAPP,并调整GPP和法尼基焦磷酸(Farnesyl diphosphate,FPP)的通量分布,在GPP合成途径中也具有重要的意义。华东理工大学花强团队[16]在解脂耶氏酵母产芳樟醇工程菌株中引入3个拷贝的IDI1基因,芳樟醇产量为0.75 mg/L;当共过表达HMG1和IDI1基因时,芳樟醇产量提高至1.44 mg/L,几乎是对照菌株的16倍,是单独过表达HMG1菌株的2.8倍。
天津科技大学于爱群团队[17]同样在解脂耶氏酵母产柠檬烯工程菌株中采用相似的策略,分别单基因过表达MVA通路中涉及到的十个酶的编码基因ACOAAT1、ACOAAT2、HMGS、HMGR、MK、PMK、PMVADO、IPPDI、GGPPS和FPPS,其中单基因过表达HMGR基因的菌株合成D型和L型柠檬烯的产量最高,分别达到0.256和0.316 mg/L,该研究确定了柠檬烯合成过程中的关键限速酶为HMGR。
2.1.2 关键酶的改造和调控
单萜产量的最大化由单位菌体生产能力和菌体总量决定,通过对关键酶蛋白进行直接改造和调控的方式来提高关键酶的活性可有效解除MVA途径中的关键酶限速步骤,从而提高单位菌体的生产能力。该方法也有利于避免由基因过表达对细胞所产生的代谢负担。
微生物中缺乏特异性的GPPS,在法尼基焦磷酸合酶(ERG20)催化IPP和DMAPP生成GPP的反应过程中,它会再次以GPP为底物生成FPP,这也是造成在微生物细胞中合成单萜类化合物的直接前体GPP供给不足的主要原因。华东理工大学花强团队[16]在解脂耶氏酵母初始菌株中引入ERG20F88W-N119W基因,造成了ERG20的氨基酸残基发生了突变,从而改变了GPP和FPP通量的分布,大大提高了GPP的积累量。与对照菌株相比,以改造后的工程菌株作为出发平台,其异源合成芳樟醇的产量大大提升。浙江大学叶丽丹团队[18]通过对F96W、N127W和K197G这三个突变位点进行组合诱变以进一步改变了ERG20的底物选择性,结果表明在线粒体和细胞质中表达改造型ERG20F96W-N127W基因的酿酒酵母工程菌株合成芳樟醇的产量最高,达到2.69 mg/L,与表达野生型ERG20基因的菌株相比增加了2倍。
GPPS和柠檬烯合酶(limonene synthesis,LS)的N端都含有一段靶向序列,对这段靶向序列进行的截短修饰有利于提高酶的催化活性[19]。美国劳伦斯伯克利国家实验室Taek Soon Lee团队[20]在大肠杆菌中构建出L-柠檬烯的合成途径后,通过对GPPS和LS这两种酶进行截短优化,使得L-柠檬烯的产量从2 mg/L提升到了约40 mg/L。其次,使用强启动子提高MK和PMK的表达水平,并用来自金黄色葡萄球菌的HMGS和HMGR来替换大肠杆菌的内源酶,L-柠檬烯的产量从2 mg/L提升到了70 mg/L。最后,结合使用上述两种策略,L-柠檬烯的产量从2 mg/L又提升到了335 mg/L。
天津大学元英进团队[2]在改造酿酒酵母菌株生产香叶醇的过程中依托计算机应用软件去认识和理解关键酶的结构,通过理性设计达到了对酶进化和修饰的目的。首先基于计算机对导肽结构的预测结果,在香叶醇合酶(CrGES)N端4个不同的位置(S14、L28、S43和S52)分别进行截断,并通过建模分析发现在S43处去除导肽序列的香叶醇合成酶(t3CrGES)的二级结构稳定性比其余3种酶更高,结果表明t3CrGES截短型菌株合成香叶醇的产量最高,达到191.61 mg/L,是未截短型菌株的4.45倍,说明了导肽和蛋白质结构稳定性对酶活性有重大影响。之后在蛋白质相互作用理论的指导下,用短柔性连接体GSG(微蛋白支架)分别从正向和反向两个角度来连接法尼基焦磷酸合酶ERG20F96W-N127W和t3CrGES以产生融合蛋白质,使得两种酶表现出更高的催化活性并减少了中间产物的损耗,结果表明组合型为t3CrGES-ERG20F96W-N127W+ERG20F96W-N127W的菌株合成香叶醇的产量最高,达到523.96 mg/L,体现出在合成途径中缩短多个酶的空间距离,能够有效地提高酶的催化反应效率。该实验结果体现出了酶工程作为关键方法改造微生物底盘合成单萜类产物的重要性,对进一步提高香叶醇产量很有指导意义。
2.1.3 区室化工程
在酵母亚细胞域细胞质内采用组合共表达MVA途径中的途径基因、突变关键酶ERG20的基因位点,以及对关键酶进行融合表达(如利用蛋白支架连接)等方法目前已经成为了代谢工程改造微生物异源合成单萜芳香产品研究中比较常态化的策略,而把异源途径转移到微生物其它亚细胞区室中的报道却很少看到。浙江大学叶丽丹团队[18]在酿酒酵母不同亚细胞区室(如线粒体和细胞质)中构建了双重MVA代谢途径,在双向GAL1/GAL10启动子的作用下,以单顺反子形式在线粒体和细胞质中同时表达芳樟醇合酶(linalool synthase,LIS)基因和ERG20F96W-N127W基因,合成芳樟醇的产量达到7.61 mg/L,与仅在细胞质中表达MVA途径相比,芳樟醇的产量得到明显提升,这种区室化工程策略为今后芳樟醇和其它单萜类化合物的高水平生产提供了较好的思路。
2.2 发酵优化策略
根据微生物在生长繁殖过程中对外界环境的要求,对微生物的发酵条件和营养条件进行优化,能够有效提高其细胞生长速率和代谢产物合成速率,进而实现利用微生物生产更多特定代谢产物的目标。在经代谢工程改造后能够异源合成单萜芳香产品的菌株中,原有的代谢途径及调控机制已被改变,此时的发酵优化就显得尤为重要。
2.2.1 优化发酵条件
天津科技大学于爱群团队[17]利用代谢工程技术改造解脂耶氏酵母异源生产柠檬烯后,设计实验验证了镁离子对解脂耶氏酵母工程菌株合成柠檬烯的促进作用,并利用单因素实验法(温度、pH、转速、初始细胞密度、添加剂)对工程菌株进行发酵优化试验,确定了最佳的工艺条件:发酵温度20 ℃,转速250 r/min,初始OD600=2.0,pH 5.74和培养基体积50 mL(250 mL摇瓶中),正十二烷体积10%,培养时间5 d,MgSO4·7H2O浓度0.2%。实验过程中获得的D-柠檬烯和L-柠檬烯的最高产量分别达到11.705 和11.088 mg/L。
中科院青岛生物能源与过程研究所咸谟和刘会洲团队[21]从优化发酵培养方式和培养环境入手来提升大肠杆菌工程菌株的生产性能,进而提高香叶醇的产量:a.对大肠杆菌工程菌株进行摇瓶发酵培养(OD600=2),48 h后产量达到68.6 mg/L;另外对该菌株进行分批补料发酵培养,IPTG诱导5 h,产量提高到78.8 mg/L。b. 在没有葡萄糖的情况下,大肠杆菌细胞会重复利用醋酸盐,从而促进乙酰基酯酶(Acetylesterase,Aes)催化乙酸香叶酯转化生成香叶醇。于是,通过对大肠杆菌工程菌株采用葡萄糖饥饿策略(在48 h停止葡萄糖供应,在葡萄糖不足的条件下继续培养)进行培养,分批补料发酵后成功将浓度为1.27 g/L(88.8%)的乙酸香叶酯转化为香叶醇,香叶醇的终产量达到2.0 g/L。
2.2.2 优化培养基碳源
华东理工大学花强团队[16]利用解脂耶氏酵母产芳樟醇工程菌株来探究葡萄糖、甘油、果糖或柠檬酸(各20 g/L)分别作为单一碳源对细胞生长和合成芳樟醇的影响,结果表明柠檬酸盐组合成芳樟醇的产量最高,产量和细胞干重(dry cell weight,DCW)分别达到2.52 mg/L和356 μg/g。另外还探究了10 g/L柠檬酸盐与10 g/L葡萄糖、甘油或果糖的混合碳源对细胞生长和合成芳樟醇的影响,观察到柠檬酸盐和葡萄糖作为混合碳源的细胞生长情况最好,而芳樟醇的产量与以20 g/L柠檬酸盐作为唯一碳源组相比几乎保持不变。最后,工程菌株在以20 g/L柠檬酸盐和8 g/L丙酮酸盐为碳源时进行摇瓶培养合成芳樟醇的产量最高,达到了6.96 mg/L(939 μg/g DCW)。
在此基础上,华东理工大学花强团队[22]继续对解脂耶氏酵母工程菌株生产柠檬烯的发酵条件进行了优化,首先探究了8种初始浓度为20 g/L的碳源(甘油、葡萄糖、柠檬酸、果糖、麦芽糖、蔗糖、甘露糖和半乳糖)对解脂耶氏酵母合成柠檬烯的影响,结果表明以甘油为碳源时合成柠檬烯的产量最高,达到1.74 mg/g DCW。再通过在培养基中添加不同初始浓度(10、20、30、40、50 g/L)的甘油来探究碳源浓度对解脂耶氏酵母生产柠檬烯的影响,结果表明,初始浓度为20 g/L的甘油组合成柠檬烯的产量最高,达到1.47 mg/g DCW;并观察到随着甘油初始浓度的升高,柠檬烯产量达到峰值的时间点会出现向后推迟的现象。此外,该团队还探究了不同初始浓度(0~4 g/L)的柠檬酸盐、乙酸盐、丙酮酸盐和苹果酸盐作为辅助碳源对解脂耶氏酵母生产柠檬烯的影响情况,结果表明初始浓度为4 g/L的柠檬酸盐组作为辅助碳源时合成柠檬烯的产量最高,达到58.4 mg/L,比对照组高1.39倍。最后该团队以甘油作为主要碳源,并补充辅助碳源柠檬酸,在1.5 L发酵罐中进行分批补料发酵培养,柠檬烯的终产量达到了165.3 mg/L,这也是目前为止解脂耶氏酵母异源合成柠檬烯的最高产量。
浙江大学叶丽丹团队[18]同样采用分批发酵培养方式,将酿酒酵母产芳樟醇工程菌株在尿嘧啶缺陷型培养基中进行培养,并对辅助碳源进行了优化。结果表明补充辅助碳源对提高细胞生产芳樟醇的性能有很大:添加丙酮酸(4 g/L)组芳樟醇的最高产量达到21.01 mg/L,添加甲羟戊酸内酯(70 mg/L)组芳樟醇的最高产量达到23.45 mg/L。
众所周知,不同菌体对原料的利用能力不同,各种原料的利用效率之间也相差很大,并且原材料组分的差异导致各种类型培养基的价格相去甚远,因此在满足微生物生长生产要求的前提下,应该优先选择价格低、效果好的培养基。天津科技大学于爱群团队[17]探究了不同添加量(0、10%、30%、50%和70%)的厨房废油替代葡萄糖为唯一碳源对解脂耶氏酵母工程菌株生产柠檬烯的影响,结果表明:工程菌株在添加浓度为70%的厨房废油的培养基中发酵效果最好,D型和L型柠檬烯的产量分别达到2.514 和2.723 mg/L,比初始产量提高了20倍,这一研究成果也为今后利用微生物转化廉价碳源合成高附加值产品的研究带来了希望。
2.3 产物分离方法
随着微生物细胞中生化反应的进行,微生物生长环境中代谢产物的浓度会不断升高,从而对细胞产生毒害或者代谢负担[24-25],最终导致单萜产品的产量和细胞生物量之间呈现反比例关系。在微生物发酵工程中,对目标产物进行有效、及时地分离可有助于解决该问题。
中国科学院青岛生物能源与过程研究所咸谟和刘会洲团队[21]对大肠杆菌产香叶醇工程菌株进行分批补料发酵培养时发现,向生产系统中加入香叶醇标准品,在发酵过程的前5个小时内,生产系统因挥发作用损失了81.4%的香叶醇。该团队还发现向生产系统中添加十四酸异丙酯形成的水、有机两相系统可有效防止目标产物的挥发,也同时降低了目标产物对细胞的毒性,产物产量大大提升。
荷兰瓦赫宁根大学植物生理学实验室的Harro Bouwmeester团队和荷兰瓦赫宁根国际植物研究中心的Jules Beekwilder团队[23]利用酵母菌ERG20基因突变株AE9K197G探索在培养系统中(1.7 g/L酵母氮基(不含氨基酸)、5 g/L(NH4)2SO4、20 g/L D-葡萄糖、20 g/L琼脂)捕获目标产物柠檬烯的有效手段,比较了包括戊烷萃取、固相微萃取、十二烷覆盖萃取、特殊吸收器吸收在内的四种不同的捕获方法,证明了在微生物发酵系统中同步采取捕获产品的措施可有效地将柠檬烯提取出来,并且获得的产品与基于植物的提取系统相比更加稳定、也达到了食品级的要求。该研究成果也展现了利用微生物合成法生产挥发性单萜芳香产品的优势。
美国劳伦斯伯克利国家实验室Taek Soon Lee团队[20]以大肠杆菌产紫苏醇工程菌株为研究对象,探究了基于阴离子交换树脂的原位产品回收方法对目标产物紫苏醇生产的影响,发现了树脂Amberlite IRA 410 Cl(A)能够特异性捕获紫苏醇,也最大限度地提高了工程菌株的生产能力,使得紫苏醇的终产量达到105 mg/L。但该树脂的添加也会引起细胞毒性或机械应力从而降低细胞密度。
3. 结论与展望
综上所述,本文从代谢工程改造思路、发酵优化策略及产物分离方法等角度出发综述了利用代谢工程技术改造不同微生物宿主合成天然单萜芳香产品的研究进展。从目前报道的结果来看,利用代谢工程改造策略来优化或改变微生物宿主已有的代谢和表达调控网络,有助于大大提高目标产物产量。总之,通过代谢工程手段所构建的能够合成单萜芳香产品的微生物细胞工厂为这一重要植物天然产物的绿色生产提供了一条新路线。
但是,目前利用代谢工程技术改造微生物直接发酵生产各类单萜芳香产品的研究还面临诸多挑战,主要包括:a. 所用微生物底盘的不同对目的产物的终产量是有明显影响的,而目前的研究还主要局限于大肠杆菌、酿酒酵母和解脂耶氏酵母中(表1);b. 相关产物在这几种微生物底盘中的产量基本都是毫克级(表1),还很难满足产业化生产的实际要求。因此,在这一背景下,为了构建更高效合成单萜芳香产品的微生物细胞工厂并最终实现其绿色制造,亟需深入探究影响不同微生物底盘高效生产特定目标产物的关键机制及核心问题。要进一步明确不同微生物底盘中合成单萜芳香产品的关键控制节点和与之相对应的控制策略,应从以下几个层面入手:
3.1 培养基方面
众所周知,培养基是由人工配制的营养基质,是微生物各种生命活动的物质基础。不同微生物底盘对各种培养基原料的利用能力和效率之间相差很大,因此培养基成分是决定微生物底盘合成目标产物(尤其是异源产物)能力高低的重要因素之一。例如,之前的研究已经证实了在YPD培养基中添加适量镁离子能够提高解脂耶氏酵母D-柠檬烯合酶(d-limonene synthase,DLS)和L-柠檬烯合酶(l-limonene synthase,LLS)的催化活性同时提高柠檬烯的产量[17];在YPD培养基中补充适量丙酮酸有利于增加酿酒酵母的菌体浓度同时提高芳樟醇的产量[18]。因此,为了获得更高的目标产品产量,从培养基方面可以从进一步优化发酵培养基成分和筛选产品合成途径中相关限速酶激活剂两方面着手。
3.2 底盘细胞方面
底盘细胞是各种生化反应发生的直接场所,因此它的重要地位不言而喻。近些年,依托基因测序技术和生物信息学的高速发展,对各种微生物底盘细胞自身特征的认识已经取得了很大突破。例如,研究者对作为原核模式微生物代表的大肠杆菌的生理和代谢特征了解最为充分,应用最为广泛,其生产的许多转基因产品也已经取得了商业化。虽然在包涵体形成以及蛋白翻译后修饰等方面存在缺陷,但凭借遗传背景清晰、易培养、遗传操作性强、异源蛋白产量高等特点,大肠杆菌往往成为代谢工程中底盘细胞的首选[26-27]。对于单萜芳香产品来说,相比酿酒酵母和解脂耶氏酵母,目前大肠杆菌底盘中相关产物(柠檬烯和香叶醇)的产量也是更高的,显示出很好的应用前景。
与大肠杆菌相比,真核模式微生物酿酒酵母具有遗传稳定性更高(如能够在基因组中同时插入多个外源基因片段)、表达真核来源功能蛋白(如植物源细胞色素P450酶和单萜合酶)的能力更强[28]等特点。此外,酿酒酵母本身就具有催化产生单萜芳香产品合成所需前体物GPP的MVA途径。因此,酿酒酵母成为了异源合成单萜芳香产品的理想微生物底盘。
解脂耶氏酵母是一种新型的非模式微生物底盘细胞。相比于酿酒酵母,它具有更适合高密度发酵、碳源谱范围更广泛以及对恶劣生长环境的适应度更高等特点,使得该酵母在工业领域具有很大的应用前景[29-30]。另外,解脂耶氏酵母已经具有了MVA途径,且作为产油酵母其胞内乙酰辅酶A的积累量较高;因此在生产单萜芳香产品方面具有明显优势,应用前景同样广阔。然而由于对解脂耶氏酵母的研究及应用起步较晚,因此目前尚未完全掌握该底盘细胞内的基因表达调控等分子机制,遗传操作系统也迄待完善。
总之,目前利用代谢工程技术生产各类单萜芳香产品的研究还主要集中在大肠杆菌、酿酒酵母和解脂耶氏酵母这几种微生物底盘中。值得关注的是,依托基因测序技术和生物信息学的高速发展,近年来对许多非模式微生物底盘细胞(如蓝细菌[31-32]等)特征的认识已经取得了很大的突破,再加上CRISPR/Cas9等基因编辑技术的不断完善,有利于研究者为单萜芳香产品的微生物合成发掘新的、更有工业应用前景的非模式微生物底盘细胞,并找到不同产物与微生物底盘之间的最大兼容性。另外,利用合成生物学技术构建微生物基因组精简优化的底盘细胞[33]也有望成为提高单萜芳香产品产量的重要策略。
3.3 代谢工程策略方面
从之前的研究结果来看,利用适宜的代谢工程策略来增大代谢网络中通向目标产物合成途径的代谢流有利于提高单萜芳香产品的产量。但是,目前仍然存在很多挑战,比如:a. MVA途径自身的代谢通量过低;b. MVA途径中存在有多个分支代谢途径;c.中间体和产物的积累会造成严重的代谢负担以及毒害作用;d.对某些微生物底盘生理和代谢机制的认知还不够充分;e.无法真正意义上对微生物底盘的代谢网络进行动态调控和实现目标代谢通量的最大化。因此,很多研究者认为可以把单萜芳香产品代谢工程的优化工作分成上、中、下游三个模块来分别进行[34]:a.增加起始物质——乙酰辅酶A、丙酮酸和3-磷酸甘油的供应量[35-37];b.强化直接前体物GPP合成途径的代谢流[38-39];c.敲除或弱化直接前体物GPP的竞争性途径[18]。
目前的研究结果表明,GPP的供应不足可能是造成单萜芳香产品合成量不高的主要限速瓶颈,因此实现GPP的充足供应是提高目标产品产量的重中之重。另外,单萜芳香产品合成后存在的内源性转化现象也是导致产物产量无法大幅度提升的重要因素[23]。另外,研究者需要利用更理性全面的策略(如系统生物学)和更先进的技术和方法(如合成生物学)来深入探究影响不同微生物底盘高效生产特定目标产物的限速步骤、关键机制及核心问题。只有明确了关键限制因素,才能找准目标、有的放矢,最终实现目标产物产量、产率的大幅度提高。
3.4 代谢产物耐受性方面
随着产物的不断积累,微生物生长环境中单萜芳香产品的浓度不断升高,就产生了对微生物底盘细胞的毒害作用。单萜芳香产品对微生物细胞造成毒害的机制非常复杂,目前已知的主要原因是单萜物质引起了微生物细胞壁、细胞膜和细胞器膜等结构的破坏[40-41];降低了细胞内某些酶的活性;并阻碍了细胞生理活动的正常进行,最终导致微生物死亡。以下策略可以提高微生物底盘对代谢产物的耐受性问题:a.利用诱变育种或适应性进化等技术筛选出耐受性更强的菌株作为出发的底盘细胞;b.首先利用转录组学或蛋白质组学等方法探索微生物耐受单萜物质的机制,然后采用分子育种手段(比如引入特异性转运蛋白)来提高微生物底盘细胞对单萜产物的耐受性;c.采用萃取、树脂吸收或膜过滤等分离技术及时地把单萜产物从培养环境中分离出来,实现“边产生边分离”,阻止毒害现象的发生。
最后,利用构建多种微生物底盘细胞组成的混合发酵系统[42-43]以及无细胞表达系统[44]等策略来突破现有的一些限制性因素,从而提高代谢工程设计和改造的效率,也有助于提高利用微生物细胞工厂合成单萜芳香产品的生产水平。
-
![]()
图 1 微生物单萜及其衍生物的生物合成途径
Figure 1. Overview of the biosynthetic pathway of monoterpenes and their derivatives in microorganisms
表 1 微生物单萜及其衍生物的产量和相关的途径改造策略
Table 1 The production of monoterpenes and the related engineering strategies in microorganisms
宿主 产品 最高产量(mg/L) 改造策略和发酵方式 参考文献 解脂耶氏酵母 柠檬烯 165.3 1)增加tNDPS1和tLS基因拷贝;
2)发酵条件优化;
3)分批补料发酵。[22] 解脂耶氏酵母 柠檬烯 D型11.705 1)单基因过表达基因HMGR;
2)发酵条件优化;
3)厨房废油作为单一碳源;
4)摇瓶培养。[17] L型11.088 大肠杆菌 柠檬烯 L型435 1)单质粒异源表达多个基因;
2)提高酶MK和PMK的含量;
3)表达外源基因HMGS和HMG1和截断型GPPS酶;
4)摇瓶培养。[20] 酿酒酵母 柠檬烯 D型0.12 1)戊烷萃取;
2)固相微萃取;
3)十二烷覆盖;
4)顶空捕捉。[23] L型0.49 酿酒酵母 香叶醇 1680 1)过表达基因tHMG1和IDI1;
2)利用计算机结构分析和建模来截短CrGES氮端的转运肽;
3)将GES和Erg20偶联为融合蛋白表达;
4)分批补料发酵。[2] 大肠杆菌 香叶醇 2000 1)添加十四酸异丙酯形成水-有机两相培养体系;
2)过表达乙酰酯酶Aes基因,验证了大肠杆菌能够水解乙酸香叶酯;
3)采用葡萄糖饥饿策略;
4)摇瓶培养。[21] 大肠杆菌 紫苏醇 105 1)诱导剂IPTG和阿拉伯糖诱导细胞来提高P450表达水平;
2)Amberlite树脂回收产品;
3)摇瓶培养。[20] 解脂耶氏酵母 芳樟醇 6.96 1)共过表达HMG1、IDI1、ERG20F88W-N119W三个基因;
2)添加混合碳源、辅助碳源来优化培养基;
3)摇瓶培养。[16] 酿酒酵母 芳樟醇 23.45 1)突变基因ERG20的K197位点;
2)线粒体区室化;
3)添加辅助碳源来优化培养基;
4)分批发酵。[18]  下载: 导出CSV
下载: 导出CSV
-
[1] Aguilar S R, Stülke J, Dijl J M V. Less is more: Toward a genome-reduced Bacillus cell factory for ‘Difficult proteins’[J]. ACS Synthetic Biology,2019,8(1):99−108. doi: 10.1021/acssynbio.8b00342
[2] Jiang G Z, Yao M D, Wang Y, et al. Manipulation of GES and ERG20 for geraniol overproduction in Saccharomyces cerevisiae[J]. Metabolic Engineering,2017,41:57−66. doi: 10.1016/j.ymben.2017.03.005
[3] Wu J, Cheng S, Cao J, et al. Systematic optimization of limonene production in engineered Escherichia coli[J]. Journal of Agricultural and Food Chemistry,2019,67(25):7087−7097. doi: 10.1021/acs.jafc.9b01427
[4] Ciriminna R, Lomeli-Rodriguez M, Demma Carà P, et al. Limonene: A versatile chemical of the bioeconomy[J]. Chemical Communications,2014,50(97):15288−15296. doi: 10.1039/C4CC06147K
[5] Tracy N I, Chen D C, Crunkleton D W, et al. Hydrogenated monoterpenes as diesel fuel additives[J]. Fuel,2009,88(11):2238−2240. doi: 10.1016/j.fuel.2009.02.002
[6] 任建峰, 张其胜, 王慧. 紫苏醇对结肠癌SW480细胞凋亡的影响及其机制研究[J]. 肿瘤药学,2019,9(3):396−400. [7] Arunachalam K, Ramar M, Ramanathan S, et al. In vivo protective effect of geraniol on colonization of Staphylococcusepidermidis in rat jugular vein catheter model[J]. Pathogens and Disease,2018,76(5):fty055. doi: 10.1093/femspd/fty055
[8] Alves S, Duarte A, Sousa S, et al. Study of the major essential oil compounds of Coriandrum sativum against Acinetobacter baumannii and the effect of linalool on adhesion, biofilms and quorum sensing[J]. Biofouling,2016,32(1−2):155−165.
[9] Lee S C, Wang S Y, Li C C, et al. Anti-inflammatory effect of cinnamaldehyde and linalool from the leaf essential oil of Cinnamomum osmophloeum Kanehira in endotoxin-induced mice[J]. Journal of Food and Drug Analysis,2018,26(1):211−220. doi: 10.1016/j.jfda.2017.03.006
[10] Lee B K, Jung A N, Jung Y S. Linalool ameliorates memory loss and behavioral impairment induced by REM-Sleep deprivation through the serotonergic pathway[J]. Biomolecules Therapeutics,2018,26(4):368−373. doi: 10.4062/biomolther.2018.081
[11] 庞亚如, 胡智慧, 肖冬光, 等. 柠檬烯和红没药烯的微生物代谢工程[J]. 生物工程学报,2018,34(1):34−43. [12] Hu Z H, Tang B, Wu Q, et al. Transcriptome sequencing analysis reveals a difference in monoterpene biosynthesis between scented Lilium 'Siberia' and unscented Lilium 'Novano'[J]. Frontiers in Plant Science,2017,8:1351. doi: 10.3389/fpls.2017.01351
[13] Frank A, Groll M. The methylerythritol phosphate pathway to isoprenoids[J]. Chemical Reviews,2017,117(8):5675−5703. doi: 10.1021/acs.chemrev.6b00537
[14] Liao P, Hemmerlin A, Bach T J, et al. The potential of the mevalonate pathway for enhanced isoprenoid production[J]. Biotechnology Advances,2016,34(5):697−713. doi: 10.1016/j.biotechadv.2016.03.005
[15] Jozwiak A, Lipko A, Kania M, et al. Modeling of dolichol mass spectra isotopic envelopes as a tool to monitor isoprenoid biosynthesis[J]. Plant Physiology,2017,174(2):857−874. doi: 10.1104/pp.17.00036
[16] Cao X, Wei L J, Lin J Y, et al. Enhancing linalool production by engineering oleaginous yeast Yarrowia lipolytica[J]. Bioresource Technology,2017,245(Part B):1641−1644.
[17] Pang Y, Zhao Y, Li S L, et al. Engineering the oleaginous yeast Yarrowia lipolytica to produce limonene from waste cooking oil[J]. Biotechnology for Biofuels,2019,12(1):241. doi: 10.1186/s13068-019-1580-y
[18] Zhang Y, Wang J, Cao X, et al. High-level production of linalool by engineered Saccharomyces cerevisiae harboring dual mevalonate pathways in mitochondria and cytoplasm[J]. Enzyme and Microbial Technology,2020,134:109462. doi: 10.1016/j.enzmictec.2019.109462
[19] Burke C, Croteau R. Geranyl diphosphate synthase from Abies grandis: cDNA isolation, functional expression, and characterization[J]. Archives of Biochemistry and Biophysics,2002,405(1):130−136. doi: 10.1016/S0003-9861(02)00335-1
[20] Aonso-Gutierrez J, Chan R, Batth T S, et al. Metabolic engineering of Escherichia coli for limonene and perillyl alcohol production[J]. Metabolic Engineering,2013,19:33−41. doi: 10.1016/j.ymben.2013.05.004
[21] Liu W, Xu X, Zhang R, et al. Engineering Escherichia coli for high-yield geraniol production with biotransformation of geranyl acetate to geraniol under fed-batch culture[J]. Biotechnol for Biofuels,2016,9:58. doi: 10.1186/s13068-016-0466-5
[22] Cheng B Q, Wei L J, Lv Y B, et al. Elevating limonene production in oleaginous yeast Yarrowia lipolytica via genetic engineering of limonene biosynthesis pathway and optimization of medium composition[J]. Biotechnology and Bioprocess Engineering,2019,24(3):500−506. doi: 10.1007/s12257-018-0497-9
[23] Jongedijk E, Cankar K, Ranzijn J, et al. Capturing of the monoterpene olefin limonene produced in Saccharomyces cerevisiae[J]. Yeast,2015,32(1):159−171.
[24] Belletti N, Kamdem S S, Tabanelli G, et al. Modeling of combined effects of citral, linalool and beta-pinene used against Saccharomyces cerevisiae in citrus-based beverages subjected to a mild heat treatment[J]. International Journal of Food Microbiology,2010,136(3):283−289. doi: 10.1016/j.ijfoodmicro.2009.10.030
[25] Saharan R K, Sharma S C. Effects of glutathione modulation on oxidative stress and enzymatic antioxidant defence in yeast Pachysolen tannophilus[J]. Current Microbiology,2011,62(3):944−949. doi: 10.1007/s00284-010-9808-x
[26] 王诗语, 王鹤蓉, 黄子豪, 等. 利用基因工程改造的大肠杆菌合成蒎烯[J]. 生物技术通讯,2019,30(4):455−463. doi: 10.3969/j.issn.1009-0002.2019.04.001 [27] Zada B, Wang C, Park J B, et al. Metabolic engineering of Escherichia coli for production of mixed isoprenoid alcohols and their derivatives[J]. Biotechnology for Biofuels,2018,11:210. doi: 10.1186/s13068-018-1210-0
[28] Krivoruchko A, Nielsen J. Production of natural products through metabolic engineering of Saccharomyces cerevisiae[J]. Current Opinion in Biotechnology,2015,35:7−15. doi: 10.1016/j.copbio.2014.12.004
[29] Jin C C, Zhang J L, Song H, et al. Boosting the biosynthesis of betulinic acid and related triterpenoids in Yarrowia lipolytica via multimodular metabolic engineering[J]. Microbial Cell Factories,2019,18:77. doi: 10.1186/s12934-019-1127-8
[30] Spagnuolo M, Shabbir H M, Gambill L, et al. Alternative substrate metabolism in Yarrowia lipolytica[J]. Frontiers in Microbiology,2018,9:1077. doi: 10.3389/fmicb.2018.01077
[31] Formighieri C, Melis A. A phycocyanin·phellandrene synthase fusion enhances recombinant protein expression and β-phellandrene (monoterpene) hydrocarbons production in Synechocystis (cyanobacteria)[J]. Metabolic Engineering,2015,32:116−124. doi: 10.1016/j.ymben.2015.09.010
[32] Shabestary K, Anfelt J, Ljungqvist E, et al. Targeted repression of essential genes to arrest growth and increase carbon partitioning and biofuel titers in cyanobacteria[J]. ACS Synthetic Biology,2018,7(7):1669−1675. doi: 10.1021/acssynbio.8b00056
[33] 林章凛, 张艳, 王胥, 等. 合成生物学研究进展[J]. 化工学报,2015,66(8):2863−2871. [34] Lv X, Xie W, Lu W, et al. Enhanced isoprene biosynthesis in Saccharomyces cerevisiae by engineering of the native acetyl-CoA and mevalonic acid pathways with a push-pull-restrain strategy[J]. Journal of Biotechnology,2014,186:128−136. doi: 10.1016/j.jbiotec.2014.06.024
[35] Hirokawa Y, Kubo T, Soma Y, et al. Enhancement of acetyl-CoA flux for photosynthetic chemical production by pyruvate dehydrogenase complex overexpression in Synechococcus elongatus PCC 7942[J]. Metabolic Engineering,2020,57:23−30. doi: 10.1016/j.ymben.2019.07.012
[36] Webb J P, Arnold S A, Baxter S, et al. Efficient bio-production of citramalate using an engineered Escherichia coli strain[J]. Microbiology,2018,164(2):133−141. doi: 10.1099/mic.0.000581
[37] Wadhwa M, Srinivasan S, Bachhawat AK, et al. Role of phosphate limitation and pyruvate decarboxylase in rewiring of the metabolic network for increasing flux towards isoprenoid pathway in a TATA binding protein mutant of Saccharomyces cerevisiae[J]. Microbial Cell Factories,2018,17(1):152. doi: 10.1186/s12934-018-1000-1
[38] Deng Y, Sun M, Xu S, et al. Enhanced (S)-linalool production by fusion expression of farnesyl diphosphate synthase and linalool synthase in Saccharomyces cerevisiae[J]. Journal of Applied Microbiology,2016,121(1):187−195. doi: 10.1111/jam.13105
[39] Liu G S, Li T, Zhou W, et al. The yeast peroxisome: A dynamic storage depot and subcellular factory for squalene overproduction[J]. Metabolic Engineering,2020,57:151−161. doi: 10.1016/j.ymben.2019.11.001
[40] Gu H, Zhu Y, Peng Y, et al. Physiological mechanism of improved tolerance of Saccharomyces cerevisiae to lignin-derived phenolic acids in lignocellulosic ethanol fermentation by short-term adaptation[J]. Biotechnology for Biofuels,2019,12(1):268. doi: 10.1186/s13068-019-1610-9
[41] Zhang L, Xiao W H, Wang Y, et al. Chassis and key enzymes engineering for monoterpenes production[J]. Biotechnology Advances,2017,35(8):1022−1031. doi: 10.1016/j.biotechadv.2017.09.002
[42] Weber W, Daoud-El Baba M, Fussenegger M. Synthetic ecosystems based on airborne inter- and intrakingdom communication[J]. Proceedings of the National Academy of Sciences of the United States of America,2007,104(25):10435−10440. doi: 10.1073/pnas.0701382104
[43] Minami H, Kim J S, Ikezawa N, et al. Microbial production of plant benzylisoquinoline alkaloids[J]. Proceedings of the National Academy of Sciences of the United States of America,2008,105(21):7393−7398. doi: 10.1073/pnas.0802981105
[44] Ward V C A, Chatzivasileiou A O, Stephanopoulos G. Cell free biosynthesis of isoprenoids from isopentenol[J]. Biotechnology and Bioengineering,2019,116(12):3269−3281. doi: 10.1002/bit.27146
 下载:
下载:

计量
- 文章访问数:
- HTML全文浏览量:
- PDF下载量:
